The Journal of Neuroscience, August 1, 1999, 19(15):6623–6628
Attenuation of Emotional and Nonemotional Memories after their Reactivation: Role of  Adrenergic Receptors Jean Przybyslawski, Pascal Roullet, and Susan J. Sara Neuromodulation et Processus Cognitifs, Institut des Neurosciences, Centre National de la Recherche Scientifique, Unite´Mixte de Recherche 7624, Universite´ Paris VI, 75005 Paris, France
A memory trace in its active state is susceptible to interference
when administered immediately after the initial training. These
by amnesic agents, such as hypothermia and electroconvulsive
results suggest that reactivation of memory triggers a 
shock, and by NMDA receptor antagonists, suggesting that a
receptor-dependent cascade of intracellular events, recapitu-
time-dependent consolidation process occurs each time a
lating that which occurs during initial postacquisition consoli-
memory is reactivated. The role of  noradrenergic receptors in
dation, thus permitting reorganization of the existing memory as
reconsolidation in rats was examined in both a positively rein-
a function of new information in the retrieval environment. This
forced radial maze task and a footshock-reinforced conditioned
remarkable lability of an active memory trace provides a new
emotional response task. For the former, rats were trained over
basis for pharmacotherapeutic intervention in such syndromes
several days in a spatial reference memory task and received a
as Posttraumatic Stress Disorder.  adrenoreceptor antago-
single reactivation trial followed by propranolol. A temporally
nists may be promising pharmacological agents for attenuating
graded impairment was observed when propranolol treatment
debilitating memories at the time of their controlled reactivation.
occurred after the memory reactivation trial. In the emotionaltask, memory impairing effects of propranolol were greater
Key words:  receptors; memory reactivation; propranolol;
when the drug was administered after a reactivation trial than
CREB; post-traumatic stress disorder; amnesia
Memories are reactivated by cues associated with the initial
1997). This suggests that the cascade of intracellular events in-
acquisition of information. Repeated reactivation of a memory
volved in plasticity and memory formation and dependent on
may serve to reinforce it and promote its long-term consolidation
NMDA receptor action is recapitulated each time the memory
(Squire and Alvarez, 1995). A more dynamic view holds that
memory is a reconstruction and reorganization of past experi-
The question arises as to whether other intracellular pathways
ences within the current cognitive context (Bartlett, 1932; Lewis
thought to be involved in long-term memory (LTM) formation
et al., 1972; Lewis and Bregman, 1973; Spear and Mueller, 1984;
are also involved in a reconsolidation process after memory
Sara, 1985). According to this view, each time a memory is
reactivation. The cAMP response element-binding protein
retrieved, it is integrated into ongoing perceptual and emotional
(CREB) pathway is one system receiving recent attention con-
experiences and becomes part of a new memory.
cerning its possible role in LTM (for review, see Mayford et al.,
A memory trace in its active state is vulnerable to interference
1995; Yin and Tully, 1996). Mice lacking CREB genes show
by amnesic agents, such as electroconvulsive shock (Misanin et
deficits in long-term potentiation and LTM (Bourtchuladze et al.,
al., 1968; Schneider and Sherman, 1968) or hypothermia (Riccio
1994); antisense oligodeoxynucleotides directed against CREB
and Stikes, 1969; Richardson et al., 1982). Such experiments
mRNA can inhibit LTM (Guzowski and McGaugh, 1997). The 
provided experimental evidence that reactivated memories, as
noradrenergic receptor is one of a family of receptors positively
well as newly acquired information, undergo a time-dependent
coupled to adenylcyclase-linked G-protein receptors governing
consolidation process, although they did not address the question
the cAMP cascade. Indirect evidence for involvement of this
of the neurobiological mechanisms involved. We have recently
pathway in LTM can be found in studies showing that blockade of
provided evidence that NMDA receptors are involved in these
the  adrenergic receptor by propranolol immediately after mem-
reconsolidation processes, at least in memories for tasks involving
ory acquisition can, in some circumstances, produce retrograde
spatial information. Blockade of these receptors by the noncom-
amnesia in humans (Cahill et al., 1994; Nielsen and Jensen, 1994).
petitive antagonist MK801 as late as 2 hr after reactivating the
Furthermore, facilitation of memory retrieval processes by stim-
memory produces a memory deficit (Przybyslawski and Sara,
ulation of the noradrenergic system is blocked by the  receptor
antagonist propranolol (Devauges and Sara, 1991). Finally, there
Received April 1, 1999; revised May 11, 1999; accepted May 12, 1999.
is growing evidence that hippocampal synaptic plasticity is de-
This research was supported by the European Foundation Programme on the
pendent on  receptor-mediated modulation (Harley, 1987;
Neural Mechanisms of Learning and Memory and by the Centre National de la
Recherche Scientifique, Unite´ Mixte de Recherche 7624. We thank Yves Moricard
In the present series of experiments, the role of
for help in preparing the figures and this manuscript.
Correspondence should be addressed to Susan J. Sara, Neuromodulation et
evaluated in postreactivation reconsolidation in two distinct be-
Processus Cognitifs, Institut des Neurosciences, Centre National de la Recherche
havioral situations: one a nonstressful appetitive task and the
Scientifique, Unite´ Mixte de Recherche 7624, Universite´ Paris VI, 9 quai St.
other a conditioned emotional avoidance response. A spatial
Copyright 1999 Society for Neuroscience 0270-6474/99/196623-06$05.00/0
reference memory task that draws minimally on working memory
6624 J. Neurosci., August 1, 1999, 19(15):6623–6628
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
was used as the appetitive task. As a control for the specificity of
the rats were tested under the same conditions as the reactivated group
the amnesic effect on active memory, two replication experiments
included control groups trained, but not receiving the reactiva-
To control for possible effects of propranolol on motivation to con-
sume Chocopops, a control experiment was performed using eight rats
tion trial before the drug treatment. A final study controlled for
from the nonreactivated group having served in the final reactivation
possible taste aversion induced by propranolol.
replication experiment. The rat was placed in the box in which it had the
A single trial inhibitory avoidance task was used as the aversive
initial exposure to reinforcement during the pretraining period, and the
training. The advantage of this behavioral procedure is that the
latency and time taken to consume five Chocopops were recorded. The
time of learning can be fixed with precision. The first phase of this
rat was then injected with propranolol 5 min later. Twenty-four hours
later, the latency and consumption time was again recorded under the
experiment evaluated the effects of propranolol injected after
same conditions. Data were analyzed by a paired t test comparing latency
acquisition. In the second phase of the experiment, control rats
to eat and total time to consume the ration, before and after drug
showing a robust memory (100% avoidance) after the memory
had been reactivated were injected with saline or propranolol and
Inhibitory avoidance training apparatus and procedure. The apparatus
consisted of two 18 cm cubic boxes constructed from Perspex, one white
and one black, each with a transparent cover. The white box had a
Perspex floor and was separated by a sliding door from the black box,
which had a grid floor through which a scrambled shock (0.25 mA for 2
MATERIALS AND METHODS
sec) could be delivered. Rats were placed individually in the white box of
Animals. One hundred ten naive male Sprague Dawley rats, weighing
the training apparatus facing the closed door. After 15 sec, the door was
between 250 and 300 gm, obtained from IFFA Credo (Arbresle, France),
raised, and the time to enter the black box was recorded. When the rat
were used in the three radial maze experiments, and fifty-eight were used
was completely inside the black box, the door was lowered, and the
in the avoidance experiments. They were housed in pairs in wire mesh
animal received a 2 sec shock. Vocalization and jumping were noted. The
cages (35 ϫ 20 ϫ 18 cm) and maintained on a 12 hr light/dark cycle with
rat was removed and placed in the home cage. Five minutes after the
water and food available ad libitum. They were weighed and handled
shock, rats were injected with propranolol (10 mg/kg, i.p) or an equal
daily for 1 week before the beginning of the experiment. Rats used in the
volume of saline. Rats were assigned to the control (n ϭ 38) or experi-
maze experiments were mildly food deprived to ϳ90% of freely feeding
mental (n ϭ 20) groups based on their initial latency to enter the dark
weight. Chocolate flavored crisp rice cereal (Chocopops; Kellogg’s) was
box, so as to have no group differences. For the testing phase 48 hr later,
used as reinforcement in the maze, and the rats were habituated to this
the rat was placed in the white box as done previously. After 15 sec, the
food before training. All procedures were performed according to the
door was raised and stayed open for 5 min. No shock was delivered.
policy on the use of animals in neuroscience research as established by
Latency to place two paws in the black box and latency to enter with all
four paws were recorded. The latency to place two paws was subjected to
Maze training apparatus and procedure. The training procedure and
a Student’s t test. Because most control rats avoided placing all four paws
apparatus were the same as that used in previous experiments (Przybys-
into the dark compartment, yielding a large number of ceiling values of
lawski and Sara, 1997). An eight-arm radial maze was elevated 0.6 m
300 sec, the data were transformed into class frequencies of rats avoiding
from the floor. Three of the eight arms were baited, the same three for
or not avoiding for the 300 sec test period, and a 2 test was applied.
every trial for an individual rat. The maze was surrounded by a black
Twenty-one control rats avoided the dark box for the entire 5 min
curtain, and salient items were hung on the curtains to serve as distal cues
testing period. Five minutes after the test, these rats were injected with
to aid in mapping of the environment. A radio, always situated in the
propranolol (n ϭ 11) or with NaCl (n ϭ 10). A second test was conducted
same position in the room, provided a distal auditory cue. To ensure that
48 hr later in the same conditions with the same behavioral measures.
the rats were really basing their performance on the integration of spatial
Drug treatment. DL-Propranolol obtained from Sigma (St. Louis, MO)
information provided by the distal extra maze cues, the maze was rotated
was prepared in saline at a concentration of 10 mg/ml and was injected
between successive trials during both training and the test sessions. This
intraperitoneally in a volume of 1 ml/kg. This single dose was used
procedure precluded any possibility for the individual rat to use in-
because previous experiments in our laboratory indicated the 10 mg/kg
tramaze information to find the baited arms, because the location of the
intraperitoneally in the rat has no effect on spontaneous locomotor
reinforced alleys was always determined by their relation to the distal
activity or exploratory behavior (Sara et al., 1995) and is effective in
cues in the extra maze environment. The experiment began with 2 d of
blocking noradrenergically induced increases in excitability of hippocam-
pretraining in which the reinforcement was available throughout the
pal neurons (Harley and Sara, 1992).
maze. After that, the animals were submitted to three daily trials (with a
intertrial interval of 5 min), which consisted of placing the rat on the
central platform of the maze and allowing free choice of visits to the
alleys, only three of which were baited. A ceiling time of 5 min was
imposed. The exact sequence of alleys visited was noted, as well as
Temporal gradient of efficacy of propranolol after
reference errors (visits to the nonbaited arms) and working errors
reactivation of a nonemotional spatial memory
(repeated visits). Acquisition criterion was three consecutive trials with a
Twenty-four hours after the reactivation session, control rats
The day after reaching criterion, the rat received a reactivation trial,
showed very good retention performance. On the other hand, rats
which consisted of a single run in the maze, after which it was returned
injected with propranolol for up to 2 hr after reactivation made
to its home cage until the scheduled injection time. All rats performed
more errors than at the reactivation trial, as shown in Figure 1.
Data for the test session concerning the difference in total number
Experimental design and data analyses. Rats were divided into six
of errors from reactivation to test trial were submitted to a
treatment groups to be injected with propranolol or saline 5 min, 2 hr, or
5 hr after the reactivation trial to determine a temporal gradient or
two-way ANOVA. There was a significant overall drug effect
window of efficacy of drug treatment. A retention session occurred 24 hr
(F(1,52) ϭ 5.37; p ϭ 0.024); the interaction approached signifi-
after the reactivation trial. Data were analyzed using a two-way
ANOVA, with one factor being drug treatment and the other time of
(2,52) ϭ 2.77; p ϭ 0.07]. Planned comparisons using the
Fischer least significant difference test (Winer, 1962) indicated a
injection. Planned comparisons were performed using the Fischer least
significant difference test (Winer, 1962).
significant difference between propranolol-treated and saline
Two complementary experiments replicated the effect of propranolol
groups at the 5 min delay ( p Ͻ 0.01) and a significant difference
on a reactivated memory and controlled for the specificity of the effect by
between the propranolol group treated at 5 min and 5 hr after
adding a group that was not subjected to a reactivation trial. The rats
reactivation ( p Ͻ 0.05). The group treated with propranolol at 2
were trained as in the preceding experiment, and one group was sub-
jected to a reactivation trial followed by an injection of propranolol 2 hr
hr after training had an intermediary performance, which was not
later in one replication and 5 min later in the second replication. The
significantly different from either the 5 min group or the 5 hr
control groups received an injection of propranolol in the vivarium, and
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
J. Neurosci., August 1, 1999, 19(15):6623–6628 6625
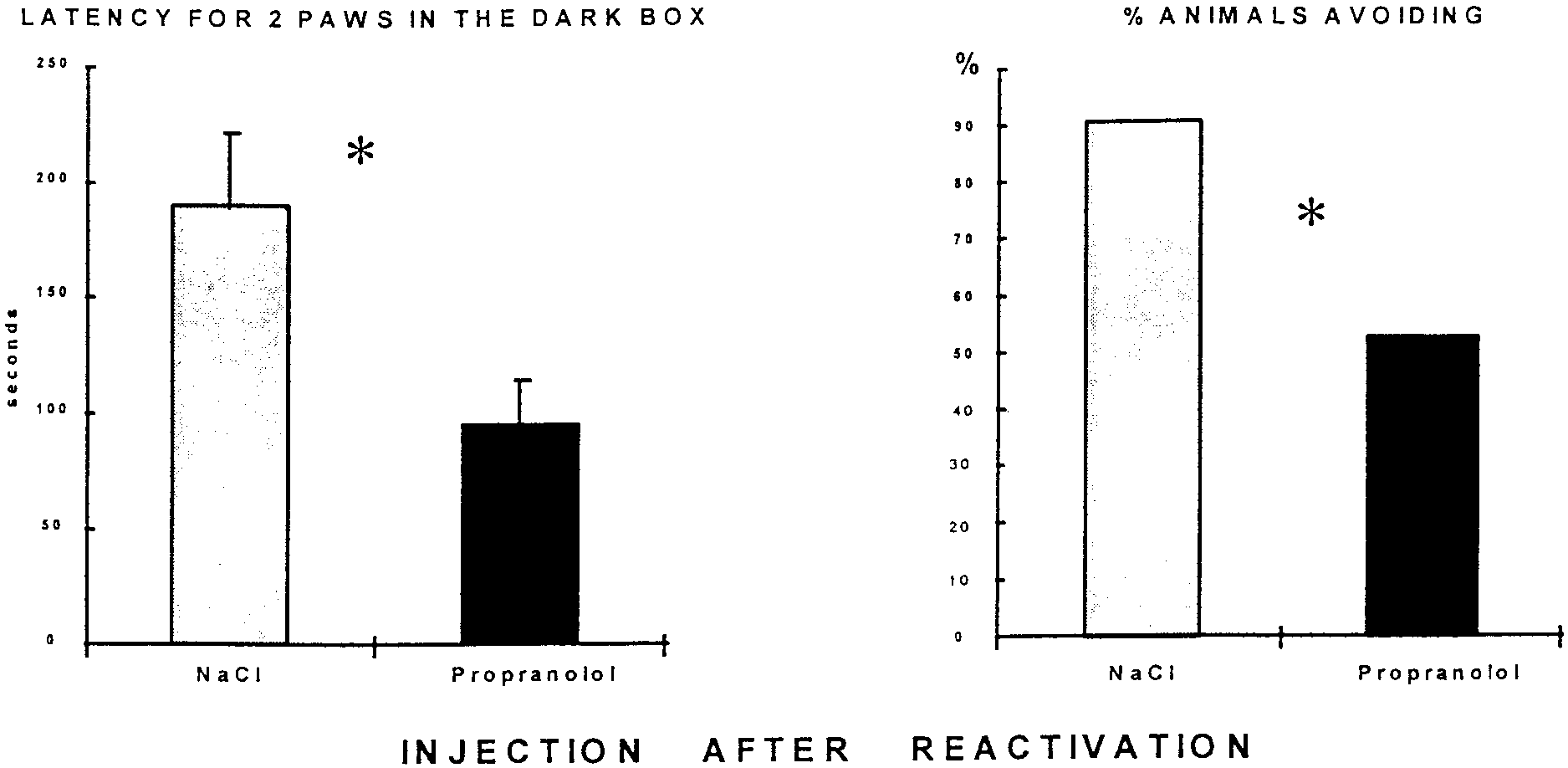
vation in the passive avoidance task caused a marked perfor-
mance decrement when the animals were retested, as shown in
Figure 5. There was a decrease in the latency to place two paws
(t(19) ϭ 2.68; p Ͻ 0.05) and in the frequency distribution of
number of rats to enter the dark box with all four paws (2 ϭ
3.231; 0.01 Ͻ p Ͻ 0.05). Note that the mean performance of the
saline control group in this phase of the experiment was better
than at the test (reactivation) trial (Fig. 4) because only those
animals treated with saline and having an optimal avoidance
performance at reactivation were used for the postreactivation
study, half being treated again with saline and half being treated
with propranolol after reactivation. DISCUSSION These experiments provide clear evidence that pharmacological
blockade of  receptors by systemic injections of propranolol up
Figure 1. Effect of propranolol at different time intervals after a reacti-
to 2 hr after reactivation of a memory trace induces amnesia
vation trial in the radial maze task. Twenty-four hours after the reacti-
when rats are tested 1 or 2 d later. The amnesia is transient or
vation session, control rats had good retention performance, whereas
propranolol-injected rats (10 mg/kg, i.p.) showed amnesia when the
partial in that the rats are capable of relearning the task with
injections were made up to 2 hr after the reactivation trial. **p Ͻ 0.01,
considerable savings. Thus far, studies of the role of  noradren-
significantly greater than saline group; ‡p Ͻ 0.05, significantly less than 5
ergic receptors in rats have suggested that these receptors play a
role in memory consolidation mainly by interaction with other
neurotransmitter systems, particularly GABAergic (Introini-
Effect of propranolol with or without reactivation
Collison et al., 1994), cholinergic (Introini-Collison et al., 1996),
Injection 5 min after a reactivation trial produced a significant
and opioid (Introini-Collison et al., 1989), the site of action being
performance decrement 24 hr later compared with the rats re-
the amygdala (for review, see McGaugh and Cahill, 1997). An
ceiving a propranolol injection in the animal vivarium, as shown
early study in adult rats did, however, show an amnesic effect of
propranolol alone when injected 5 min after passive avoidance
(14) ϭ 4.015; p ϭ 0.001). Propranolol injections 2 hr
after reactivation also induced some amnesia, because these an-
training. Interestingly, the animals were able to express memory
imals made more errors at the retention test than the control
for up to 6 hr after treatment, the amnesia appearing only in those
group injected without a reactivation trial. A t test revealed a
tested 1 d after training (Cohen and Hamburg, 1975), corrobo-
significant effect of treatment on the difference between number
rating later views that  receptors govern the adenylcyclase-
of errors on the last training trial and the mean of three test trials
linked cAMP cascade leading to protein synthesis-dependent
(26) ϭ 2.30; p Ͻ 0.05). Note that the last training trial was used
as a baseline performance in these experiments, because one
A recent study in humans suggests that propranolol selectively
group did not receive a reactivation trial.
attenuates memories for emotionally charged events (Cahill et al.,
In these two experiments, only rats that had the behavioral trial
1994). Another study in elderly humans reports that  blockers, at
before the propranolol treatment showed amnesia, although they
clinically antihypertensive doses, can block the beneficial effects
had been subjected to one more trial than the nonreactivated rats.
of arousal on memory performance (Nielsen and Jensen, 1994). It
Thus, the effect of a propranolol injection was limited to a period
should be noted that, aside from these two studies, there are few
of up to 2 hr after memory comes to an active state.
reports of memory impairment associated with clinical doses of 
blockers, despite their widespread use as antihypertensives. This
Effect of propranolol on reward incentive
is probably because of the use of hydrophilic forms, which do not
There were no differences in the latencies or total time to con-
readily cross the blood–brain barrier.
sume the Chocopops before or after propranolol treatment, as
Systemic injections, such as those used in the present experi-
indicated in Figure 3. (paired t tests; latency, t(7) ϭ 1.3; total time,
ments, are useful if the drug treatment might have a clinical
t(7) ϭ 1.36; p Ͼ 0.05).
application, as discussed below. However, this leaves open the
Propranolol injection after inhibitory avoidance training
question of site of drug action. There are compelling arguments
Propranolol-treated rats had a shorter latency to place two paws
for the effect being mediated by blockade of  receptors in the
CNS. Systemically administered propranolol has little or no effect
(56) ϭ 2.19; p Ͻ 0.05), as shown in Figure 4.
There was no effect of treatment on percentage of rats completely
on cerebral blood flow to account for its memory impairing effects
entering the dark box (2 ϭ 0.82, NS), as illustrated in Figure 4.
(Olesen, 1986). Peripherally administered propranolol, a li-
Thus, propranolol has only a small effect on memory, as measured
pophilic molecule, readily crosses the blood–brain barrier; after
by the immediate step into response. The two other behavioral
chronic treatment in humans, the ratio of brain/plasma concen-
measures most often used to evaluate memory in the passive
trations of the drug is ϳ20:1 (Cruickshank et al., 1980; Neil-
avoidance task, step-through latency and total time spent in the
Dwyer et al., 1981). An important recent study has provided
shock compartment, were not modified by post-training propran-
strong evidence that the central effects of propranolol are respon-
sible for the amnesia for emotional events in man by comparing
the effects of propranolol with a hydrophilic  antagonist, which
Propranolol after reactivation of avoidance training
does not cross the blood–brain barrier. The latter had no
Saline-treated animals continued to show good avoidance behav-
memory-impairing effects. Finally, in a recent study from our
ior at the second test. Propranolol injected 5 min after a reacti-
laboratory, intracerebroventricular injection of the  antagonist
6626 J. Neurosci., August 1, 1999, 19(15):6623–6628
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
Figure 2. Effect of propranolol on memory with or without reactivation trial. A, Rats received a propranolol injection (10 mg/kg, i.p.) in the animal
vivarium 2 hr after a reactivation trial or after no reactivation trial (n ϭ 14 in each group). There was a significant performance deficit in the group that
had the reactivation trial before the drug treatment compared with the group that received drug alone. *p Ͻ 0.05. B, Procedure the same as in A, except
that injections were made 5 min after a reactivation trial or in the vivarium (n ϭ 8 in each group). There was a significant performance deficit in rats
receiving the injection after a reactivation trial compared with those rats receiving drug treatment without reactivation. Note the nearly errorless
performance of this group on the test trial. **p Ͻ 0.001.
of amnesic agents (Misanin et al., 1968; Schneider and Sherman,1968; Riccio and Stikes, 1969; Lewis et al., 1972; Lewis andBregman, 1973). The results of those early experiments, althoughthey did not extend our knowledge of the neurobiological pro-cesses underlying these reconsolidation processes, did reinforcethe notion that memory is dynamic and that new memories areformed on the foundation of reactivated old memories. Thatpostreactivation amnesia can be induced by both NMDA recep-tor (Przybyslawski and Sara, 1997) and  receptor blockade
(Roullet and Sara, 1998) suggests intracellular mechanisms in-volving the same second messenger pathways as involved in syn-aptic plasticity and initial memory formation.
In the present series of experiments, the effect of propranolol is
not limited to conditioned emotional responses but can be ob-tained in appetitive situations in which the animals are only mildlyfood-deprived and are well trained in the task. Two controlprocedures ensured that the behavioral deficit was not caused bya proactive effect of propranolol on performance at the time oftest. Those animals injected 5 hr after reactivation and tested 24hr later showed no such performance decrement nor did thosethat were not subjected to a reactivation trial before drug treat-ment. Memory must be in an active state for propranolol to beeffective, and the temporal limit for treatment efficacy under
Figure 3. Effect of propranolol on reward incentive. Latency (left) and
total time (right) to consume five Chocopops before (white bars) and 24 hr
these experimental conditions is between 2 and 5 hr.
after (black bars) injection of propranolol. There is no significant change
It is surprising that the effect of propranolol appeared to be
in either measure (paired t test), indicating that the drug did not induce
more robust after reactivation of inhibitory avoidance training
than after the original learning. Such increased vulnerability toamnesic agents after reactivation is not, however, unprecedented.
timolol impaired a reactivated memory in a paradigm similar to
Mactutus et al. (1979) reported that memory reactivated by ex-
the one used in the present experiments. Intracerebroventricular
posure to the place where a footshock had been administered was
injections allowed a temporal resolution of drug action and a
more susceptible to hypothermia-induced amnesia than immedi-
window of efficacy was found at 1 hr after the reactivation trial;
ately after the initial acquisition. Further investigation is required
intracerebroventricular injections earlier or later did not impair
to determine whether reactivated memories, in general, are more
labile and vulnerable to amnesic agents or whether it is particular
The present results reinforce previous studies showing that
to the conditioned emotional response elicited by the passive
reactivated memories are susceptible to interference by a variety
avoidance test. It is possible that the increased vulnerability to the
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
J. Neurosci., August 1, 1999, 19(15):6623–6628 6627 Figure 4. Effect of propranolol injection 5 min after passive avoidance training. Left, Latency to place two paws into the dark box in the test session
48 hr after training. Propranolol (10 mg/kg, i.p.) significantly decreased this measure of retention. Right, Percentage of rats completely entering the dark
box; there was no difference between control (n ϭ 38) and propranolol-injected (n ϭ 20) rats on this measure of retention. Figure 5. Effect of propranolol 5 min after a reactivation in passive avoidance task. Left, Latency to place two paws in the shock compartment in the
test session 48 hr after reactivation session. Propranolol-injected rats (10 mg/kg, i.p.) show a significant decrease in latency. Right, Percentage of rats
completely entering the dark box. There was a significant increase in the propranolol-treated group on this measure of retention (control, n ϭ 10;
propranolol, n ϭ 11; *p Ͻ 0.05). Note that only those rats treated with saline and showing perfect retention at the reactivation phase were used in this
phase of the experiment, which accounts for the improvement in performance in saline-treated group shown here compared with Figure 4. Half were
treated again with saline and half received propranolol after the reactivation trial.
amnesic agent after the retention–reactivation test is because the
a possible clinical application in the pharmacotherapeutic treat-
rat receives no footshock during the exposure. Some extinction
ment of Posttraumatic Stress Disorder (PTSD). This psychiatric
could be occurring, although this is not seen in the subsequent
syndrome is characterized by vivid recall of the traumatic events
behavior of the saline-injected control group, who maintain max-
with the accompanying severe emotional responses. Individuals
imal avoidance behavior at the second retention test. We are
report that terrifying experiences are often recalled with inten-
currently developing rapidly learned appetitive tasks in our lab-
sity, the traumatic events being reexperienced unchanged over
oratory to perform comparative studies between emotional and
years (van der Kolk and Fisler, 1995). There is rather extensive
nonemotional memories and effects of reactivation procedures
evidence that points to dysregulation of the noradrenergic system
in PTSD, and there has been some suggestion of how this might
This demonstration of lability of reactivated memories suggests
be related to the hypermnesia. Over-responsiveness of the nor-
6628 J. Neurosci., August 1, 1999, 19(15):6623–6628
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
adrenergic system during stress could recreate the internal state
-noradrenergic influences on memory storage involve cholinergic ac-
induced by the original trauma and thereby “reinstate” the mem-
tivation. Neurobiol Learn Mem 65:57–64.
ory (Grillon et al., 1996). The potential usefulness of noradren-
Lewis D, Bregman NJ, Mahan J (1972) Cue-dependent amnesia in rats.
J Comp Physiol Psychol 81:243–247.
ergic receptor-blocking agents in PTSD has already been pointed
Lewis DJ, Bregman NJ (1973) Source of cues for cue-dependent amne-
out by Cahill (1997), who suggests that treatment with  blockers
sia in rats. J Comp Physiol Psychol 85:421–426.
as soon as possible after the traumatic event might prevent the
Mactutus CF, Riccio DC, Ferek JM (1979) Retrograde amnesia for old
development of PTSD. The lability of memory in its active state,
(reactiviated) memory: some anomalous characteristics. Science
demonstrated in the present experiments, adds a new dimension
Mayford M, Wang J, Kandel ER, O’Dell TJ (1995) CaMKII regulates
to this potential use. Effective treatment might lie in reactivation
the frequency-response function of hippocampal synapses for the pro-
of the traumatic memory under psychotherapeutic conditions
duction of both LTD and LTP. Cell 81:891–904.
combined with pharmacological treatment with a  receptor
McGaugh JL, Cahill L (1997) Interaction of neuromodulatory systems in
antagonist to reduce the strength of the memory. The results of
modulating memory storage. Behav Brain Res 83:31–38.
the present experiments suggest that treatment with propranolol,
Misanin JR, Miller RR, Lewis DJ (1968) Retrograde amnesia produced
by electroconvulsive shock after reactivation of consolidated memory
especially at the time of spontaneous or clinically elicited rein-
statement of the traumatic memory, should serve to attenuate the
Neil-Dwyer G, Bartlett J, McAinsh J, Cruickshank JM (1981)
active memory by blocking reconsolidation processes.
-Adrenoreceptor and the blood–brain barrier. Br J Clin Pharmacol
REFERENCES
Nielson KA, Jensen RA (1994) -Adrenergic receptor antagonist anti-
Bartlett FC (1932) Remembering. A study in experimental and social
hypertensive medications impair arousal-induced modulation of work-
psychology. Cambridge, MA: Cambridge UP.
ing memory in elderly humans. Behav Neural Biol 62:190–200.
Bourtchuladze R, Frenguelli B, Blendy J, Cioffi D, Schutz G, Silva AJ
Olesen J (1986) -Adrenergic effects on cerebral circulation. Cephalal-
(1994) Deficient long-term memory in mice with a targeted mutation
of the cAMP-responsive element-binding protein. Cell 79:59–68.
Przybyslawski J, Sara SJ (1997) Reconsolidation of memory after its
Cahill L (1997) The neurobiology of emotionally influenced memory.
reactivation. Behav Brain Res 84:241–246.
Implications for understanding traumatic memory. Ann NY Acad Sci
Riccio DC, Stikes ER (1969) Persistent but modifiable retrograde am-
nesia produced by hypothermia. Physiol Behav 4:649–652.
Cahill L, Prins B, Weber M, McGaugh JL (1994)  adrenergic activation
Richardson R, Riccio DC, Mowrey H (1982) Retrograde amnesia for
and memory for emotional events. Nature 371:702–704.
previously acquired pavlovian conditioning: UCS exposure as a reac-
Cohen RP, Hamburg MD (1975) Evidence for adrenergic neurons in a
tivation treatment. Physiol Psychol 10:384–390.
memory access pathway. Pharmacol Biochem Behav 3:519–523.
Roullet P, Sara SJ (1998) Consolidation of memory after its reactivation:
Cruickshank JM, Neil-Dwyer G, Cameron MM, McAinsh J (1980)
involvement of  noradrenergic receptors in the late phase. Neural
␣-Adrenoreceptor-blocking agents and the blood–brain barrier. Clin
Sara SJ (1985) Noradrenergic modulation of selective attention: its role
Devauges V, Sara SJ (1991) Memory retrieval enhancement by locus
in memory retrieval. Ann NY Acad Sci 444:178–193.
coeruleus stimulation: evidence for mediation by beta receptors. Behav
Sara SJ, Dyon-Laurent C, Herve´ A (1995) Novelty seeking behavior in
the rat is dependent upon the integrity of the noradrenergic system.
Grillon C, Southwick SM, Charney DS (1996) The psychobiological ba-
sis of posttraumatic stress disorder. Mol Psychiatry 1:278–297.
Sara SJ, Roullet P, Przybyslawski J (1999) Consolidation of memory for
Guzowski JF, McGaugh JL (1997) Antisense oligodeoxynucleotide-
odor-reward association:  adrenergic receptor involvement in the late
mediated disruption of hippocampal cAMP response element binding
protein levels impairs consolidation of memory for water maze training.
Sarvey JM (1988) Protein synthesis in long-term potentiation and
Proc Natl Acad Sci USA 94:2693–2698.
norepinephrine-induced long-lasting potentiation in hippocampus. In:
Harley CW (1987) A role for norepinephrine in arousal, emotion and
Long-term potentiation: from biophysics to behavior (Liss AR, ed), pp
learning?: limbic modulation by norepinephrine and the Kety hypoth-
esis. Prog Neuropsychopharmacol Biol Psychiatry 11:419–458.
Schneider AM, Sherman W (1968) Amnesia: a function of the temporal
Harley CW, Sara SJ (1992) Locus coeruleus bursts induced by glutamate
relation of footshock to electroconvulsive shock. Science 159:219–221.
trigger delayed perforant path spike amplitude potentiation in the
Spear NE, Mueller CW (1984) Consolidation as a function of retrieval.
dentate gyrus. Exp Brain Res 89:581–587.
In: Memory consolidation: psychobiology of cognition (Weingartner H,
Huang, YY, Kandel ER (1996) Modulation of both the early and the late
Parker ES, eds). Hillsdale, NJ: Erlbaum.
phase of mossy fiber LTP by the activation of -adrenergic receptors.
Squire LR, Alvarez P (1995) Retrograde amnesia and memory consoli-
dation: a neurobiological perspective. Curr Opin Neurobiol 5:169–177.
Introini-Collison IB, Nagahara AH, McGaugh JL (1989) Memory en-
van der Kolk BA, Fisler R (1995) Dissociation and the fragmentary
hancement with intra-amygdala post-training naxolone is blocked by
nature of traumatic memories: overview and exploratory study.
concurrent administration of propranolol. Brain Res 476:94–101.
Introini-Collison IB, Castellano C, McGaugh JL (1994) Interaction of
Winer BJ (1962) Statistical principles in experimental design New York:
gabaergic and -noradrenergic drugs in the regulation of memory
storage. Behav Neural Biol 61:150–155.
Yin JCP, Tully T (1996) CREB and the formation of long-term memory.
Introini-Collison IB, Dalmaz C, McGaugh JL (1996) Amygdala
Understanding Indigestion and Ulcers Professor C.J. Hawkey and Dr N.J.D. WightPublished by Family Doctor Publications Limitedin association with the British Medical Association IMPORTANT This book is intended not as a substitute for personalmedical advice but as a supplement to that advice for the patient who wishes to understand more about his In particular (without limit) you should no
ALGORITHMS FOR NONORTHOGONAL APPROXIMATE JOINTInstitute of Information Theory and AutomationPod vod´arenskou vˇeˇz´ı 4, 18208 Prague 8, Czech RepublicApproximate joint block diagonalization (AJBD) of a set ofmatrices has applications in blind source separation, e.g., n W T , when the signal mixtures contain mutually independent sub-are all approximately block diagonal, having the bl
 Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
J. Neurosci., August 1, 1999, 19(15):6623–6628 6625
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
J. Neurosci., August 1, 1999, 19(15):6623–6628 6625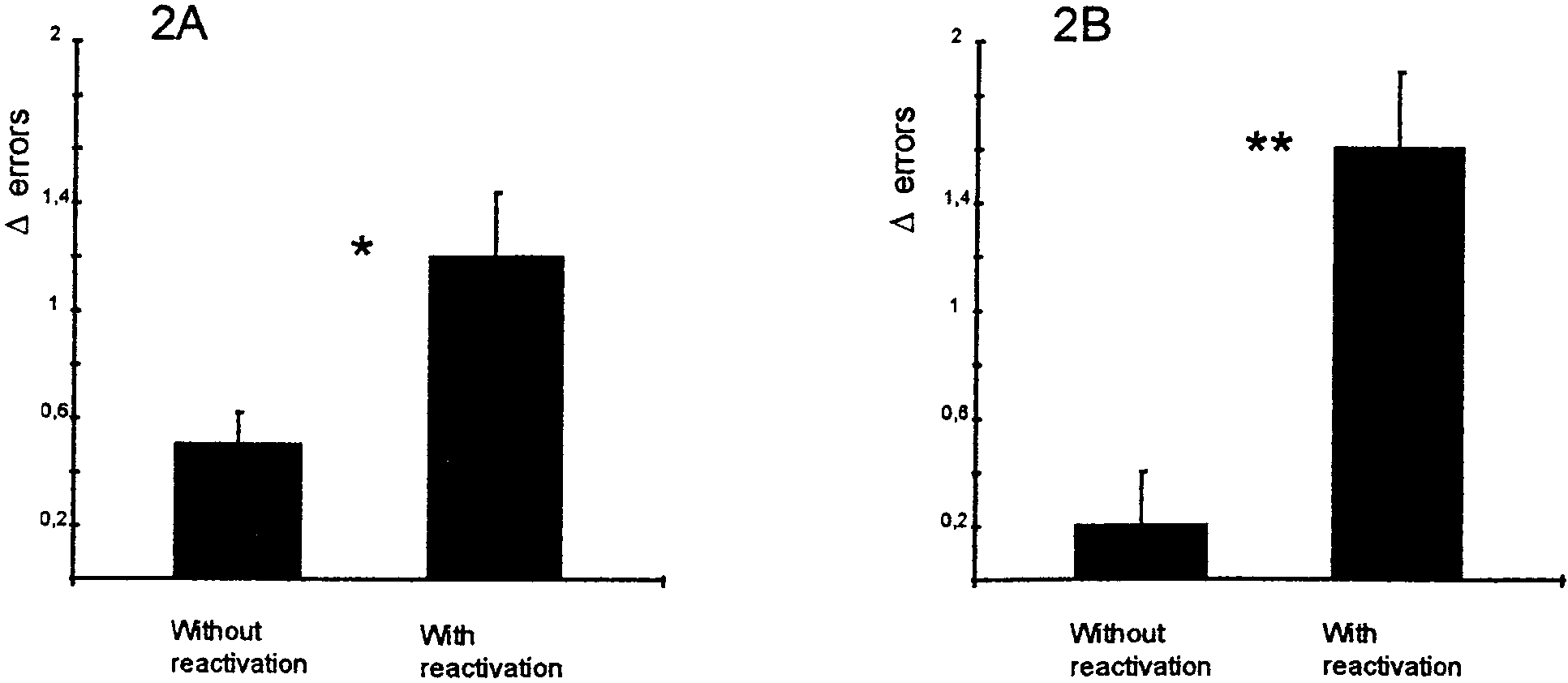
 6626 J. Neurosci., August 1, 1999, 19(15):6623–6628
6626 J. Neurosci., August 1, 1999, 19(15):6623–6628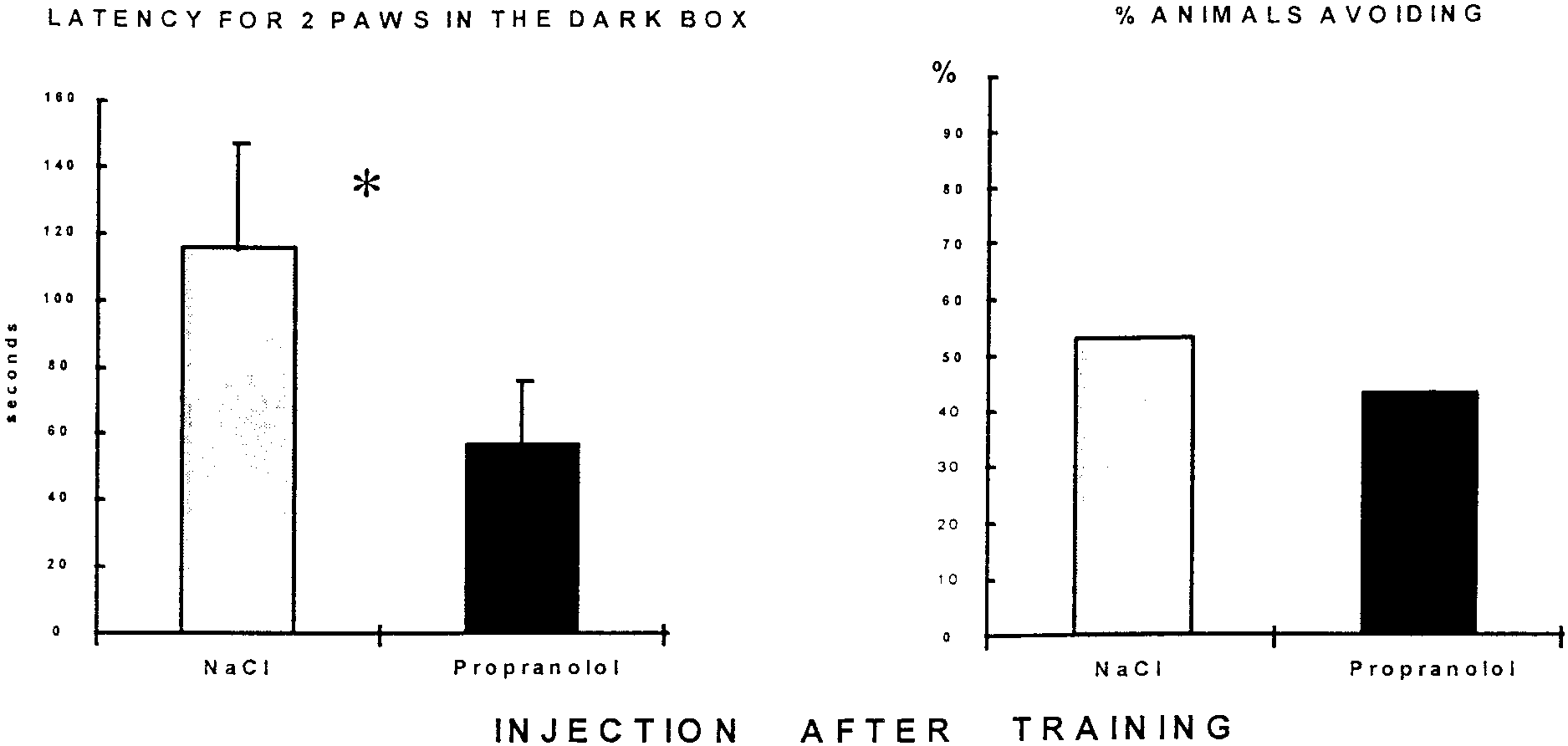
 Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
J. Neurosci., August 1, 1999, 19(15):6623–6628 6627
Przybyslawski et al. •  Noradrenergic Blockade Attenuates Reactivated Memory
J. Neurosci., August 1, 1999, 19(15):6623–6628 6627