tional specification of ventricular myosin
inducible expression in cultured neonatal
light chain 2 expression in the primitive
murine heart tube. Proc Natl Acad Sci USA
Zou Y, Chien KR: 1995. EFIA/YB-1 is a com-
Zammit PS, Kelly RG, Franco D, et al.: 2000.
and positively regulates transcription of
Suppression of atrial myosin gene expres-
the myosin light chain-2 v gene. Mol Cell
ventricular specificity and establishes an
sion occurs independently in the left and
anterior-posterior gradient of expression.
Zou Y, Evans S, Chen J, et al.: 1997. CARP, a
Seidman CE, Schmidt EV, Seidman JG: 2001.
Zeller R, Bloch KD, Williams BS, et al.: 1987.
Cis-dominance of rat atrial natriuretic fac-
Localized expression of the atrial natriuret-
tor gene regulatory sequences in transgen-
ic factor gene during cardiac embryogene-
Zhu H, Garcia S, Ross RS, et al.: 1991. A
Small EM, Krieg PA: 1999. Expression of at-
rial natriuretic factor (ANF) during Xeno-
the rat cardiac myosin light chain-2 gene
confers cardiac specific and a-adrenergic
Small EM, Krieg PA: 1976. Transgenic analy-
sis of the atrial natriuretic factor (ANF) pro-moter: Nkx2-5 and GATA-4 binding sites arerequired for atrial specific expression ofANF. Dev Biol 261:116 – 131.
Sucov HM, Dyson E, Gumeringer CL, et al.:
2002. RxRa mutant mice establish a geneticbasis for vitamin A signaling in heart mor-
phogenesis. Genes Dev 8:1007 – 1018.
Takimoto E, Mizuno T, Terasaki F, et al.:
2002. Up-regulation of natriuretic peptidesin the ventricle of Csx/Nkx2-5 transgenicmice. Biochem Biophys Res Commun 270:
Platelets are blood cell fragments that originate from the cytoplasm of
megakaryocytes in the bone marrow and circulate in blood to play a
major role in the hemostatic process and in thrombus formation after
an endothelial injury. Recent studies have provided insight into platelet
mRNA distribution patterns in the rat heart. Cardiovasc Res 32:886 – 900.
functions in inflammation and atherosclerosis. A range of molecules,
present on the platelet surface and/or stored in platelet granules, con-
tributes to the cross-talk of platelets with other inflammatory cells
during the vascular inflammation involved in the development and
& N Rosenthal (Eds.), Heart development( pp. 357 – 369). San Diego, CA: Academic
progression of atherosclerosis. This review discusses the nature of these
molecules and the mechanisms involved in the participation of plate-
Wang GF, Nikovits W, Schleinitz M, Stock-
lets in atherosclerosis, with emphasis on P-selectin, platelet–monocyte
dale FE: 1998. Atrial chamber-specific ex-
interactions, chemokines, and inflammatory cytokines. (Trends
pression of the slow myosin heavy chain 3gene in the embryonic heart. J Biol Chem
Cardiovasc Med 2004;14:18–22) n 2004, Elsevier Inc.
Wang GF, Nikovits W Jr, Schleinitz M, Stock-
dale FE 2001. A positive GATA element and
a negative vitamin D receptor-like element
Cardiovascular Research Center and Depart-
control atrial chamber-specific expression
ment of Biomedical Engineering, University
of Virginia, Health Science Center, Charlot-
tection of activated platelets as defined
diovascular Research Center and Department
Wang GF, Nikovits W Jr, Bao ZZ, Stockdale
FE: 2003. Irx4 forms an inhibitory complex
Virginia, Health Science Center, Box 801394,
atherosclerotic disease was first reported
Charlottesville, VA 22908, USA. Tel.: (+1) 434-
tors to regulate cardiac chamber-specific
by Fitzgerald et al. (1986). These circu-
243-9351; fax: (+1) 434-924-2828; e-mail:
lating activated platelets are very likely
to associate with thrombotic events. Cir-
D 2004, Elsevier Inc. All rights reserved.
findings did not support a beneficial ef-
fect of aspirin in atherosclerosis (Cayatte
et al. 2000, Napoli et al. 2002). Also, the
et al. 1998). Interestingly, most risk fac-
inhibitory effect of aspirin in atheroscle-
tors of atherosclerosis—including hyper-
cholesterolemia (Broijersen et al. 1998),
smooth muscle cells in the vessel wall.
vated platelets in circulation. The pres-
platelets transiently interacting with the
ble for platelet activation in atherosclero-
A23187 (Frenette et al. 1995). Consistent
platelet activation begins with the bind-
ing of adhesive receptors to their ligands
bution of platelet – endothelial inter-
but robustly with atherosclerotic carotid
facilitate the delivery of platelet-derived
et al. 2003). These interactions are main-
let – endothelial interactions in the devel-
and stimulated platelets, epinephrine re-
(Huo et al. 2003). Platelet P-selectin is
important. Repeated injections of P-selec-
phase of atherosclerosis, platelet activa-
lial ligand for platelet P-selectin is un-
mice accelerated the formation of athero-
sclerotic lesions (Huo et al. 2003). Recon-
of atherosclerosis, including superoxide,
for platelet translocation and firm adhe-
control mice (Burger and Wagner 2003).
sion, respectively. In addition to P-selec-
atherosclerosis is controversial. In pigs
ecule in both resting and activated plate-
sites of arteries of rabbits with hypercho-
1980, Nichols et al. 1998). Deficiency of
et al. 2002) also found GPIIb/IIIa-mediat-
did not show any protective role in devel-
ditionally, following formation of lesions
opment of atherosclerosis (Sramek et al.
on the vessel wall, platelet activation may
be initiated by ligation of GPIb with the
definitions of firm adhesion are likely to
influence the results. Consistent with the
Platelet activation influences the devel-
deficient in VWF have a defect in secretion
an injection of in vitro activated platelets.
bodies. Thus, endothelial P-selectin-medi-
in these VWF mutant mice (Denis et al.
luminal surfaces of atherosclerotic arter-
conflicting data exist. For example, some
or avidity of monocyte/leukocyte integ-rins via P-selectin glycoprotein ligand-1(PSGL-1) signaling or delivery of plate-let-derived proinflammatory factors(Figure 1B). Oxidative burst occurs onmonocytes in response to platelet bind-ing. These rapid platelet-mediated regu-lations may play a role in atherosclerosisby promoting monocyte recruitment(Figure 1B). Activated platelets are alsoable to cause a variety of slow reactions. Resting monocytes do not express tissuefactor, a protein involved in the initia-tion of blood coagulation and the forma-tion of atherosclerotic lesions. However,upon interaction with platelet P-selectin,
Figure 1. Mechanisms by which activated platelets participate in the development ofatherosclerosis. (A) No interactions occur between resting platelets and monocytes. (B)
transcription of tissue factor is activated.
Activated platelets promote monocyte recruitment via platelet – monocyte interactions.
Activated platelets interacting with monocytes deliver their proinflammatory factors to
monocytes. Consequently, affinity and/or avidity of monocyte integrins are upregulated and
monocytes arrest on endothelium. Additionally, monocyte – platelet aggregates may employ
platelet P-selectin to mediate aggregates to interact with endothelium. (C) Activated platelets
promote monocyte recruitment via platelet – endothelial interactions. Activated platelets
transiently interacting with endothelium may deposit their proinflammatory factors on thesurface of endothelium, causing subsequent rolling monocyte arrest. Also, platelet-derived
proinflammatory factors may infiltrate into the vessel wall, triggering vascular cell prolifer-
are primed by several different synergis-
tic activators. Exposure of monocytes toplatelet P-selectin and platelet activating
factor mobilizes the transcription factor
bound to activated GPIIb/IIIa complex.
a ligand for leukocyte Mac-1 (Simon et al.
platelet – leukocyte aggregates are not
sclerosis (Shpilberg et al. 2002). This is
Michelson et al. (2001) found that the life
crete a different set of cytokines, includ-
mice. In atherosclerotic mice, deficiency
shedding. In our in vivo study (Huo et al.
2003), 2 to 3 hours after an injection of
activated platelets, circulating platelet –
tectable and ‘‘normal’’ leukocyte subtype
tions between monocytes and platelets.
P-selectin on activated platelets initiates
their interactions with leukocytes. Among
leukocyte subtypes interacting with acti-
shed their P-selectin (Berger et al. 1998),
vated platelets, monocytes have a compet-
suggesting that most platelet – leukocyte
activated platelets (Huo et al. 2003). The
sequester into peripheral tissues or mono-
tially bind platelets, a possible basis for
cytes phagocytose platelets bound to their
the role of activated platelets in the devel-
opment of atherosclerosis, is unknown.
ly, interactions between activated plate-
lets and leukocytes will be stabilized due
to binding of leukocyte integrins to plate-
telets are able to upregulate affinity and/
ces that are contained within the a gran-
of inflammatory reactions, as do activat-
sclerotic lesions, direct evidence is need-
ed platelets. The role of activated platelets
are endocytosed from the blood plasma.
in atherosclerotic lesions is really ‘‘plate-
rosis. Platelet factor 4 (PF-4), a member
derived by limited proteolysis from plate-
let basic protein. PF-4 causes chemotaxis
lets have recently received intense inter-
Thirty years after Ross et al. (1976) pro-
cent studies (Nassar et al. 2003) provide
protein of the TNF family originally iden-
atherosclerosis, direct evidence now sup-
4 in the development of atherosclerosis.
telets truly play an important role in the
platelets in atherosclerosis, although par-
tially suggested in primary studies, have
cleaved over a period of minutes or hours,
generating a soluble but functional frag-
rived foam cells, is able to dramatically
capable of initiating various inflammato-
ment and progression of atherosclerosis.
ry responses on endothelial cells, includ-
ing production of reactive oxygen species
(Urbich et al. 2002), expression of adhe-
et al. 2003, Sachais et al. 2002). RANTES
sion molecules (e.g., vascular cell adhe-
sion molecule 1, ICAM-1, and E-selectin),
factor (Slupsky et al. 1998). In contrast to
preparation of this manuscript. This work
sented in its mature form on the platelet
arteries, suggesting that platelet-derived
Berger G, Hartwell DW, Wagner DD: 1999. P-
selectin and platelet clearance. Blood 92:
et al. 2003, von Hundelshausen et al.
Bowie EJ, Fuster V: 2000. Resistance to athe-
rosclerosis in pigs with von Willebrand’s
is a cationic polypeptide composed of two
chains (A and B) that are linked by inter-
Broijersen A, Hamsten A, Eriksson M, et al.:
2001a. Platelet activity in vivo in hyperlipo-
jor growth factor in platelets stimulating
and proliferation associated with intimal
for these factors, especially for IL-1h.
Burger PC, Wagner DD: 2001b. Platelet P-se-
lectin facilitates atherosclerotic lesion de-
to investigate whether transient interac-
Cayatte AJ, Du Y, Oliver-Krasinski J, et al.:
nist S18886 but not aspirin inhibits athero-
genesis in apo E-deficient mice: evidence
contribute to atherosclerosis. Arterioscler
Platelet microparticles, released from ac-
Celi A, Pellegrini G, Lorenzet R, et al.: 1998b.
out the progression of atherosclerosis.
P-selectin induces the expression of tissue
aggregates are a more sensitive marker of
1998. Activated platelets induce tissue fac-
in vivo platelet activation than platelet sur-
tor expression on human umbilical vein en-
Cyrus T, Sung S, Zhao L, et al.: 1995. Effect of
face P-selectin: studies in baboons, human
dothelial cells by ligation of CD40. Thromb
low-dose aspirin on vascular inflammation,
plaque stability, and atherogenesis in low-
Sramek A, Reiber JH, Gerrits WB, Rosendaal
density lipoprotein receptor-deficient mice.
clinically relevant effect on atherogenesis:
et al.: 1993. Inhibition of lung inflammatory
reactions in rats by an anti-human IL-8 anti-
ditary bleeding tendency. Circulation 104:
DD: 1995. Defect in regulated secretion of
P-selectin affects leukocyte recruitment invon Willebrand factor-deficient mice. Proc
Napoli C, Ackah E, De Nigris, et al.: 2002.
Chronic treatment with nitric oxide-releas-
ing aspirin reduces plasma low-density lip-
platelet-endothelial cell adhesion in postca-
Fitzgerald DJ, Roy L, Catella F, Fitzgerald GA:
pillary venules. Arterioscler Thromb Vasc
1996. Platelet activation in unstable coro-
arterial oxidation-specific epitopes, and
nary disease. N Engl J Med 315:983 – 989.
Theilmeier G, Michiels C, Spaepen E, et al.:
Frenette PS, Johnson RC, Hynes MR, Wagner
mice. Proc Natl Acad Sci 99:12,467 – 12,470.
2002. Endothelial von Willebrand factor re-
DD: 1994. Platelets roll on stimulated endo-
Nassar T, Sachais BS, Akkawi S, et al.: 2003.
cruits platelets to atherosclerosis-prone
thelium in vivo: an interaction mediated by
Platelet factor 4 enhances the binding of
sites in response to hypercholesterolemia.
endothelial P-selectin. Proc Natl Acad Sci
oxidized low-density lipoprotein to vascular
wall cells. J Biol Chem 278: 6187 – 6193.
Tsao PS, Theilmeier G, Singer AH, et al.:
Furman MI, Benoit SE, Barnard MR, et al.:
Nichols TC, Bellinger DA, Reddick RL, et al.:
1994. L-arginine attenuates platelet reactiv-
1988. Increased platelet reactivity and cir-
1998. von Willebrand factor does not influ-
ity in hypercholesterolemic rabbits. Arte-
ence atherogenesis in arteries subjected to
patients with stable coronary artery disease.
altered shear stress. Arterioscler Thromb
Urbich C, Dernbach E, Aicher A, et al.: 2002.
CD40 ligand inhibits endothelial cell migra-
Hawrylowicz CM, Santoro SA, Platt FM, Un-
Nityanand S, Pande I, Bajpai VK, et al.: 1993.
tion by increasing production of endothelial
anue ER: 1986. Activated platelets express
Platelets in essential hypertension. Thromb
reactive oxygen species. Circulation 106:
IL-1 activity. J Immunol 143:4015 – 4018.
Henn V, Slupsky JR, Grafe M, et al.: 2002.
Nowak J, Murray JJ, Oates JA, Fitzgerald GA:
CD40 ligand on activated platelets triggers
1987. Biochemical evidence of a chronic ab-
et al.: 2001. Rantes deposition by platelets
normality in platelet and vascular function
Hermann A, Rauch BH, Braun M, et al.: 1992.
Platelet CD40 ligand (CD40L)—subcellular
Pratico D, Tillmann C, Zhang ZB, et al.: 2001.
Weng S, Zemany L, Standley KN, et al.: 2003.
localization, regulation of expression, and in-
h3 integrin deficiency promotes atheroscle-
hibition by clopidogrel. Platelets 12:74 – 82.
rosis and pulmonary inflammation in high-
Huo Y, Schober A, Forlow SB, et al.: 1989.
Circulating activated platelets exacerbate
mice. Proc Natl Acad Sci 98:3358 – 3363.
atherosclerosis in mice deficient in apolipo-
Ross R: 1999. Atherosclerosis—an inflamma-
Weyrich AS, McIntyre TM, McEver RP, et al.:
tory disease. N Engl J Med 340:115 – 126.
1995. Monocyte tethering by P-selectin reg-
Lindemann S, Tolley ND, Dixon DA, et al.:
Ross R, Glomset JA: 1976. The pathogenesis
ulates monocyte chemotactic protein-1 and
2002. Activated platelets mediate inflamma-
of atherosclerosis. N Engl J Med 295:369 –
tumor necrosis factor-a secretion. Signal in-
tory signaling by regulated interleukin 1be-
tegration and NF-kappaB translocation.
ta synthesis. J Cell Biol 154:485 – 490.
Ruggeri ZM: 2002. Platelets in atherothrom-
Weyrich AS, Elstad MR, McEver RP, et al.:
1994. Reduction of atherosclerosis in mice
Sachais BS, Kuo A, Nassar T, et al.: 2002.
1996. Activated platelets signal chemokine
Platelet factor 4 binds to low-density lipo-
Manduteanu I, Calb M, Lupu C, et al.: 1995.
cytic machinery, resulting in retention of
Increased adhesion of human diabetic pla-
low-density lipoprotein on the cell surface.
telets to cultured valvular endothelial cells.
J Submicrosc Cytol Pathol 24:539 – 547.
Shpilberg O, Rabi I, Schiller K, et al.: 2002.
Massberg S, Brand K, Gruner S, et al.: 1995. A
critical role of platelet adhesion in the ini-
lacking platelet glycoprotein alpha(IIb)be-
tiation of atherosclerotic lesion formation.
ta(3) (GPIIb/IIIa) and alpha(v)beta(3) recep-
tors are not protected from atherosclerosis.
Methia N, Andre P, Denis CV, et al.: 1994.
Localized reduction of atherosclerosis in
Simon DI, Chen Z, Xu H, et al.: 2000. Platelet
von Willebrand factor-deficient mice. Blood
glycoprotein Iba is a counterreceptor for
the leukocyte integrin Mac-1 (CD11b/CD18).
et al.: 1992. Circulating monocyte-platelet
Slupsky JR, Kalbas M, Willuweit A, et al.:
Dall’empirismo alle evidenze scientifiche Università degli Studi di Roma "La Sapienza" Introduzione La bronchiolite è la più comune infezione virale del e basse vie respiratorie dei bambini nel primo anno di vita, con un picco d’incidenza tra i 3 e i 9 mesi. Il quadro clinico di presentazione è il risultato del ’ostruzione su base infiammatoria del e piccole vie aere
Name __________________________________________________________________________________________ Address ________________________________________________________________________________________ City _____________________________________________ State ________________ Zip _________________ E-Mail __________________________________________________________________________________________ _______
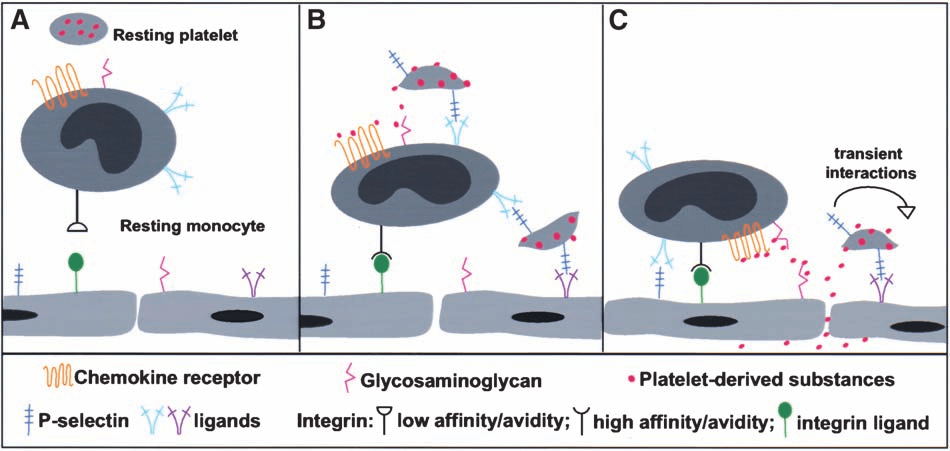 or avidity of monocyte/leukocyte integ-rins via P-selectin glycoprotein ligand-1(PSGL-1) signaling or delivery of plate-let-derived proinflammatory factors(Figure 1B). Oxidative burst occurs onmonocytes in response to platelet bind-ing. These rapid platelet-mediated regu-lations may play a role in atherosclerosisby promoting monocyte recruitment(Figure 1B). Activated platelets are alsoable to cause a variety of slow reactions.
or avidity of monocyte/leukocyte integ-rins via P-selectin glycoprotein ligand-1(PSGL-1) signaling or delivery of plate-let-derived proinflammatory factors(Figure 1B). Oxidative burst occurs onmonocytes in response to platelet bind-ing. These rapid platelet-mediated regu-lations may play a role in atherosclerosisby promoting monocyte recruitment(Figure 1B). Activated platelets are alsoable to cause a variety of slow reactions.