CURRENT MICROBIOLOGY Vol. 32 (1996), pp. 25–32
Study of the Denitrifying Enzymatic System of Comamonas sp. Strain SGLY2 Under Various Aeration Conditions with a ParticularView on Nitrate and Nitrite Reductases
Institut National de la Recherche Agronomique, Laboratoire de Biotechnologie de l’Environnement (LBE), Avenue des Etangs,11100 Narbonne, France
Abstract. This paper studies the effect of oxygen on the denitrifying enzymatic system of Comamonas sp. It is shown that nitrate respiration can take place in the presence of oxygen. Indeed, even if a protein synthesis inhibitor is added in the medium, immediate nitrate consumption is observed in an aerobic culture inoculated with cells that have never been subjected to nitrate. Existence of a constitutive nitrate reductase could explain this phenomenon. Moreover the nitrate and nitrite reductases are active and synthesized under aerobic conditions. The different levels of inhibition of nitrate reductase activity by respiratory inhibitors and detergent, according to the aerobic and anaerobic cultures, might suggest the existence of a double nitrate reductase enzymatic system.
Bacterial denitrification is an anaerobic respiration
inhibition of denitrifying enzymes with Paracoccus
where nitrate is reduced to nitrogen gas with nitrite,
denitrificans [14] or Pseudomonas aeruginosa [12];
nitric oxide, and nitrous oxide as intermediates. This
(ii) existence of a tolerance threshold towards dis-
reaction is realized by facultative anaerobic microor-
solved oxygen with Pseudomonas stutzeri; and (iii)
ganisms, especially Pseudomonas sp. [39]. It is an
co-respiration with Thiosphaera pantotropha, Alcalig-
alternative way of energy production by transfer of
enes faecalis [28, 30], Pseudomonas nautica [6] and
electrons to four de novo-synthesized terminal oxido-
reductases: nitrate reductase (NaR), nitrite reductase
Berks and associates [4, 5] have characterized the
(NiR), nitric oxide reductase (NoR), and nitrous oxide
aerobic denitrifying pathway of Thiosphaera pantotro-
reductase (N2OR). Synthesis and activity of these
pha, even though Thomsen and colleagues [37] have
enzymes are assumed to be completely repressed by
assumed that aerobic denitrification with this strain is a
oxygen and to be stimulated by one or the other of the
utopic idea because of the existence of anaerobic
microzones in the kind of reactor used. They have
More recently, some authors have demonstrated
purified a periplasmic nitrate reductase that is overex-
that synthesis and activity of denitrifying enzymes
pressed in a membrane-bound nitrate reductase mutant
could occur under various aeration conditions. For
strain [3]. They have explained the simultaneous use of
example, Pseudomonas stutzeri exhibits its higher
oxygen and nitrate by existence of this double nitrate
nitrate reductase activity at 1 mg · L21 of dissolved
reductase enzymatic system: the membrane-bound
oxygen, but the threshold value for its nitrate reductase
nitrate reductase, inhibited by oxygen, allows aerobic
synthesis is 5 mg · L21 of dissolved oxygen [15].
expression of the periplasmic one [25]. Thiosphaera pantotropha denitrifies at a rate of 800
A strain named SGLY2, and identified as Comamo-
nmol · min21 · mg21 of protein at 80% of air saturation
nas sp., has been isolated in our laboratory from an
[29]. Three different types of behavior can thus be
upflow filter submitted to various aeration conditions.
distinguished in the presence of oxygen: (i) complete
This strain was shown to denitrify in the presence ofhigh oxygen levels and to be able to co-respire the two
electron acceptors [22]. In the present study, we
characterized the enzymatic system of the strain (espe-
protein synthesis. Cells were harvested, washed, and concentrated
cially nitrate and nitrite reductases) by investigating
in 0.9% NaCl solution. They were then used to inoculate the
the influence of different respiratory protein synthesis
different flasks (control and test flasks) in anaerobic, partial aerobic,and fully aerobic conditions to reach a concentration of 35 mg · L21
inhibitors and detergent (by modifying membrane
of proteins. Flasks were shaken in a rotary shaker (200 rpm) at 35°C
permeability) on the oxygen-nitrate respiration sys-
tem. These experiments were carried out either in
Assays of nitrate and nitrite reductase activities. Nitrate and
batch culture under various aeration conditions or in
nitrite reductase activities were assayed on whole cells harvested
enzymatic assays on whole cells and cell lysates.
from three different cultures: one 400-ml aerobic culture in a 1-LErlenmeyer flask filled with the medium described above except
Materials and Methods
N-oxides; one 400-ml aerobic culture in a 1-L Erlenmeyer flaskfilled with the medium described above supplied with KNO
Organisms and culture conditions. Isolation and characterization
one 400-ml anaerobic culture in Penicillin flasks filled with the
of the strain SGLY2 used in this study have been described in detailelsewhere [22]. Cells were grown on a synthetic medium: phos-
medium supplied with KNO3. These three cultures were inoculated
(1/200) and were then grown for 16 h. After addition of chloram-
M, pH 7.0; KNO3 (N-NO3 5 250 mg · L21) or
phenicol (150 µg · ml21), cells were harvested, washed twice with
(N-NO2 5 50 mg · L21); ethanol (C-C2H5OH 5 500
0.9% cold NaCl, and suspended in the same solution.
4 190 mg · L21; (NH4)2SO4 as nitrogen source
Nitrate and nitrite reductase activities were measured accord-
4 5 58 mg · L21); yeast extract (Difco) 250 mg · L21; 1 ml ·
L21 of trace element solution [22]. To study the influence of
ing to the combined methods of Brons and Zehnder [7] and Ko¨rner
inhibitors or detergent on the oxygen-denitrifying respiration
and Zumft [15]. A 4-ml mixture consisting of 2 ml 0.1 M phosphate
system of the strain, batch cultures were performed under three
buffer, pH 7.2; 1 ml 0.1 M KNO3 or 1.5 ml 0.1 M KNO2; 0.4 ml 0.5 M
aeration conditions in 120-ml penicillin flasks (Poly Labo, Montpel-
sodium acetate and demineralized water was pipetted into 15-ml
lier), filled with the medium described above. Anaerobic conditions
tubes. The tubes were then flushed with argon and hermetically
were obtained by bubbling cultures with oxygen-nitrogen-free
sealed with rubber stoppers for anaerobic tests. Different inhibitors
were added to the reaction mixture: sodium azide to a final
CO 1 CO2 , 0.1 ppm). To obtain partial aerobic conditions, we
concentration of 100 µM and 10 mM, and erythromycin to a final
first bubbled the cultures with argon. A known quantity of pure
concentration of 200 µg · ml21. After equilibration at 35°C in a
oxygen (oxygen C Alphagas, N2 , 5 ppm) was then added into the
water bath, the reaction was started by injection of the equivalent of
sealed bottle, with a syringe, until it reached a concentration of 20
2.4 mg of cell protein per tube. For the next hour, a sample was
mmol · L21 of gas. This addition of a large quantity of oxygen
withdrawn every 20 min. The reaction was stopped by eliminating
pressurized the flasks. Since no oxygen was present in the liquid
the cells by centrifugation at 4°C. Nitrate reductase activity was
phase (because of sparging with argon), a part of the oxygen gas
expressed as mmol of nitrate consumed per minute per milligram of
was transferred to the liquid phase. At the beginning of the
proteins. Nitrite reductase activity was expressed as nmol of nitrite
experiment, this dissolved oxygen concentration corresponded to
consumed per minute per milligram of proteins.
oxygen saturation (7.8 mg · L21 at sea level at 35°C). It subse-
Nitrate reduction was also measured in cell-free extract,
quently dropped owing to the bacterial oxygen consumption,
obtained from a sonicated aerobic cell suspension, according to the
compensated by redistribution between the gaseous and liquid
modified procedure described by Krul and Veeningen [16]. The
phase. Aerobic cultures were grown in cotton-wool plugs flasks
assay mixture consisted of 2.5 ml 0.1 M phosphate buffer, pH 7.2; 1
filled with 50 ml of the medium described above, whereas in the
ml 0.1 M KNO3; 1.25 ml benzylviologen 0.2 mg · L21; and 0.25 ml
two other conditions the final volume was 100 ml. The smaller
of demineralized water. The reaction vials were sparged with argon
liquid volume in aerobic culture allowed better gas exchange.
and sealed with rubber stoppers to keep the anaerobic conditions. Inhibitors and detergent tests. Two inhibitors of cytochrome
Thereafter, 0.5 ml of a mixed solution of 10 mg · L21 Na2S2O4 and
3) at two different concentrations were tested by
3 (vol/vol) was supplied. After 15 min of
addition to the medium of sodium azide at 0.1 mM and 10 mM,
shaking in a waterbath at 35°C, the equivalent of 100 mg · L21 of
potassium cyanide at 10 and 100 µM. Diethyldithiocarbamic acid
protein extract was injected. A sample was withdrawn every 10 min
(DDC), a copper chelator, was used as an inhibitor of copper type
in a period of 30 min. The reaction was stopped by aerating the
nitrite reductase or other copper proteins of the respiratory chain
samples in order to oxidize the residual electron donor.
(azurin) at a final concentration of 10 mM. Erythromycin andchloramphenicol were used at 200 µg · ml21 and 150 µg · ml21
Analysis of biomass, medium, and gas. Nitrate and nitrite were
respectively to inhibit protein synthesis. Effect of membrane
measured by an exchange ion chromatography system with conduc-
perturbations on reductases activities was observed based on triton
tivity detection (DIONEX-100). Separation and elution of the
anions were carried out on an IonPacAS4A Analytical Column with
Inoculum was grown aerobically in a 15-ml cotton-wool plugs
a carbonate-bicarbonate eluant and a sulfuric acid regenerant.
Erlenmeyer flask containing yeast extract (5 g · L21) and peptone
Gas composition was analyzed by gas chromatography with a
(15 g · L21). After overnight growth, this culture was used to
Shimadzu GC-8A apparatus with argon carrier, by use of a
inoculate two different precultures: one 500-ml Erlenmeyer flask
katharometer detector. Carbon dioxide and nitrous oxide were
filled with 200 ml of the medium described above except N-oxides
separated on a Haye Sep Q column (80–100 mesh, 2.0 m 3 1/8
(this culture was named ‘‘nonadapted to N-oxides preculture’’) and
inch). Oxygen and nitrogen were separated on a molecular sieve 5A
one Penicillin flask filled with 200 ml of the complete medium and
(20–100 mesh, 2.0 m 3 1/8 inch). Injector and detector temperature
maintained under anaerobic conditions (this culture was named
was 100°C; column temperature was 35°C. Nitric oxide was
‘‘adapted to N-oxides preculture’’). When the precultures were in
measured on a Shimadzu-14A with helium carrier, by use of a
the logarithmic growth phase, chloramphenicol was added to stop
katharometer detector. The molecular sieve 5A (80–100 mesh,
D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.
Fig. 1. Influence of chloramphenicol (150 µg · ml21) on the nitriteproduction during anaerobic (M, N) and partial aerobic cultures(U, V) on nitrate with Comamonas sp. Cultures were inoculatedwith nonadapted preculture. Chloramphenicol was added at thebeginning of the culture. M, U, control cultures; N, V, cultureswith chloramphenicol.
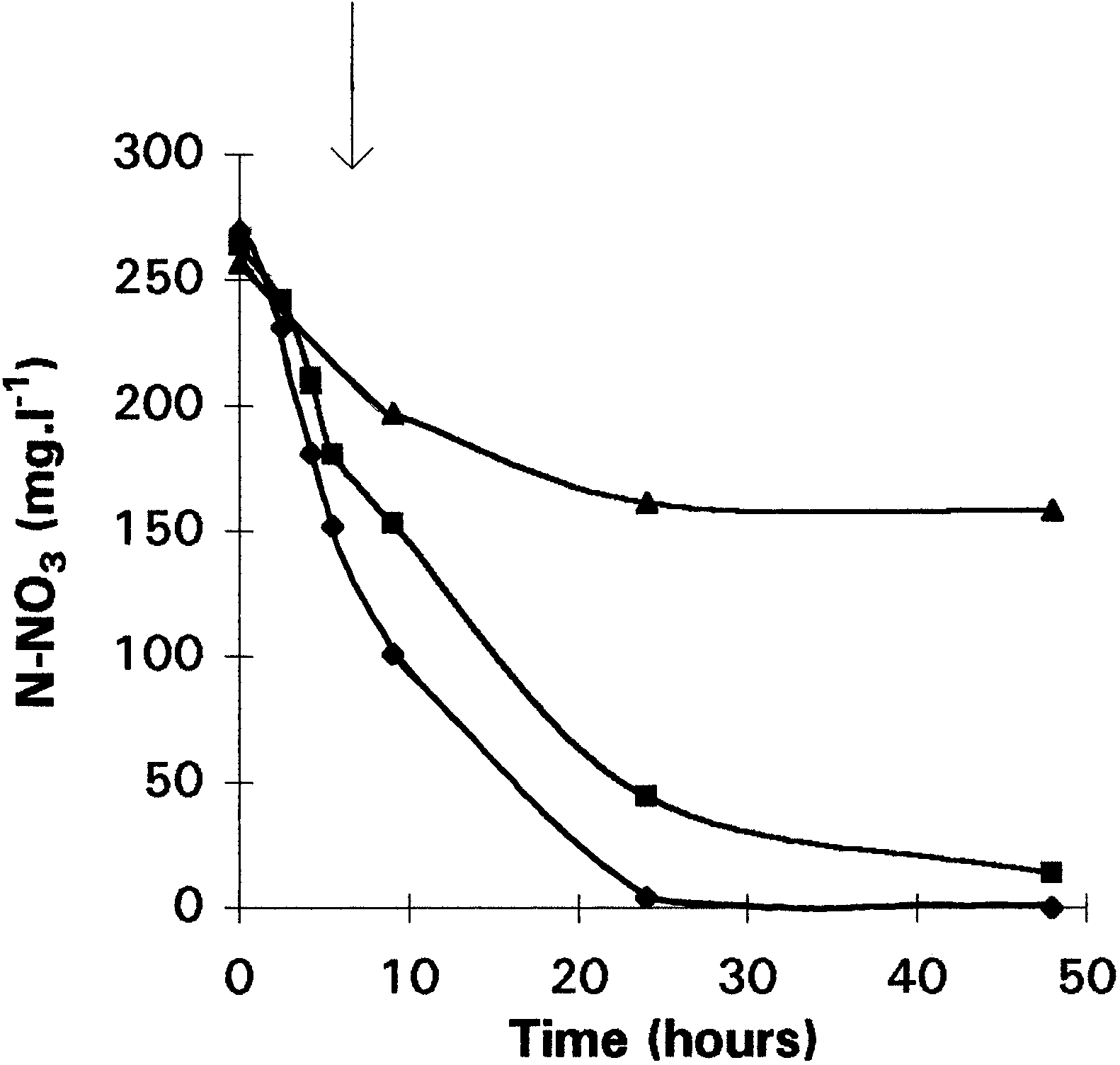
Fig. 2. Consumption of nitrate during anaerobic (M), partial aerobic(U), and total aerobic (Q) growth conditions of Comamonas sp. The arrow indicates the time at which oxygen has completelydisappeared in the partial aerobic culture. Cultures were inoculated
2.0 m 3 1/8 inch) column was maintained at 220°C, injector at
Proteins were determined by Lowry’s procedure with bovine
whereas 99 nmol were obtained in the control tube. This measurement of residual activity might be owing
to the existence of a nitrate reductase in the aerobicculture. Existence of a constitutive nitrate reductase. Chlor-
Our experiments showed that a constitutive nitrate
amphenicol (150 µg · ml21) was used to determine
reductase is present in an aerobic culture nonadapted
whether or not denitrifying enzymes were synthesized;
to N-oxides. This enzyme could be active under
nitrate reductase activities were measured in aerobic
anaerobic as well as aerobic conditions at a small basal
cultures inoculated with cells nonadapted to N-oxides.
It was added during cell harvesting by centrifugation(existence of anaerobic conditions in the pellet could
Synthesis and activity of nitrate and nitrite reducta-
enhance synthesis of enzymes) and in the different
ses under aerobic condition. The previous experi-
batch assays. Nitrate reduction was measured by
ments showed that a higher nitrate reduction rate is
production of nitrite (Fig. 1). Addition of chlorampheni-
observed when protein synthesis is not inhibited (Fig.
col in the anaerobic and partial aerobic cultures
1). In the same way, nitrate consumption began
resulted in a sharp decrease of the nitrate reduction
immediately after inoculation of the medium with
rate: 4 µg N-NO2 · h21 · mg21 of protein against 85.3
nonadapted cells, in both aerobic and anaerobic condi-
µg · h21 · mg21 of protein in anaerobic conditions.
tions (Fig. 2). During aerobic culture, nitrate consump-
However, an immediate production of nitrite was
tion slowed down after 20 h because of a lack of
noticed even though the cells used to inoculate the
carbon source. Nitrate reductase activity resulted from
culture were for the first time in the presence of nitrate.
the activity of the constitutive nitrate reductase and of
Nitrate reductase activity was also measured on
new enzyme synthesis. Thus, it seems that synthesis
SGLY2 cell extracts obtained from a culture never
and activity of nitrate reductase may occur in a wide
subjected to nitrate. In this type of culture, an activity
The same experiment was realized with nitrite as
measured. In the same way, an enzymatic assay on
the final electron acceptor (Fig. 3). Anaerobic nitrite
whole cells from aerobic culture without nitrate was
consumption started after a lag period of 1 day. In this
realized by adding erythromycin (200 µg · ml21) and
case, the use of nitrite is the only way to produce
chloramphenicol (150 µg · ml21) in the test tube. These
energy. However, according to the literature, no nitrite
protein synthesis inhibitors decreased the nitrate reduc-
reductase is synthesized in a nonadapted preculture.
tase activity: the activity, with chloramphenicol, was
This lag phase corresponds to synthesis of a de novo
nitrite reductase by using presumably residual energy
Fig. 3. Consumption of nitrite during anaerobic (M), partial aerobic
Fig. 4. Influence of triton (0.02%) on the consumption of nitrate
(U), and total aerobic cultures (Q) of Comamonas sp. The arrow
during anaerobic (M, N) and total aerobic (Q, S) cultures of
indicates the time at which oxygen has completely disappeared in
Comamonas sp. Cultures were inoculated with anaerobic cells. M,
the partial aerobic culture. Cultures were inoculated with non-
Q, control cultures; N, S; Cultures with triton; =, addition of
present in the cells. In contrast, partial or fully aerobicconsumption of nitrite started after a smaller lag period(4 h). This consumption was correlated with nitrousoxide and nitrogen production (data not shown). Oxygen consumption provided the energy necessary tosynthesize the enzyme. Addition of chloramphenicolresulted in inhibition of nitrite consumption no matterwhat culture conditions were used. These experimentsdemonstrated that nitrite reductase, which may not beconstitutive, is synthesized under aerobic conditionsand that nitrite and oxygen are consumed simulta-neously. Effect of triton, sodium azide, and cyanide on nitrate reductase activity. Figure 4 shows the nitrate consumption during anaerobic and aerobic batch cul-
Fig. 5. Influence of sodium azide (0.1 mM) on the consumption of
tures of SGLY2 inoculated with adapted-to-N-oxide
nitrate during anaerobic (M, N) and total aerobic (Q, S) cultures
cells in the presence of triton. Addition of this deter-
of Comamonas sp. Cultures were inoculated with nonadapted toN-oxides cells. M, Q, control cultures; N, S, cultures with azide.
gent to the culture medium resulted in a gentle
decrease in the anaerobic nitrate consumption (0.111mg N-NO3 · h21 · mg21 of protein against 0.062 mgN-NO3 · h21 · mg21 of protein). It seems, however, to
concentration tested (10 mM) neither denitrification
have no effect on aerobic nitrate reduction. The same
nor growth was observed. This means that the denitri-
results are obtained with the nonadapted to N-oxides
fying as well as the oxygen-respiring enzymatic sys-
preculture. The presence of triton X-100 perturbated
tem was completely inhibited at this high concentra-
the permeability properties of the cytoplasmic mem-
tion. In contrast, at 0.1 mM, the nitrate reduction rate of
brane. This implies that under anaerobic conditions,
the anaerobic culture, inoculated with the nonadapted
the measurement of nitrate reductase activity is corre-
cells (Fig. 5), fell to 7 µg N-NO3 · h21 · mg21 of
lated with the membrane, whereas aerobic nitrate
protein. This corresponds to a 97% inhibition of the
reduction is independent of the membrane.
denitrifying enzyme activity measured under control
The effect of azide on nitrate reductase activity
anaerobic conditions. During aerobic culture at 0.1
during batch assay followed exactly the same pattern
mM, no effect of azide on oxygen uptake was observed:
observed with triton (Fig. 5). Whatever the aeration
the disappearance of oxygen in the gaseous phase was
conditions and the preculture used, at the highest
correlated with protein synthesis. The only influence
D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.
Table 1. Influence of two concentrations (0.1 and 10 mM) ofsodium azide on nitrate reductase activity measured on whole cellsharvested from aerobic and anaerobic precultures (see Materialsand Methods for more details)
Activity is expressed in nmol of disappeared nitrate per minute permg of protein.
of this respiratory inhibitor on denitrifying enzymeswas then noticed. Compared with the nitrate reductionrate measured under aerobic control conditions, therewas a 61% inhibition of the nitrate reductase activity(Fig. 5). The same conclusions were drawn from themeasurements of nitrate reductase activity in wholecells (Table 1). The presence of 0.1 mM of azide in thetest tube implied a 100% fall of the nitrate reductaseactivity of anaerobic whole cells, whereas a 53%decrease was observed on nitrate reductase activity ofaerobic whole cells. The pattern of inhibition of nitrate
Fig. 6. Evolution of nitrate (M, N) and nitrite (Q, S) during
reductase activity was similar to the results obtained in
anaerobic (A) and partial aerobic (B) cultures of Comamonas sp. in
batch assays with the highest concentration of azide.
the presence of cyanide (N, S) at a final concentration of 10 µM. Cultures were inoculated with nonadapted to N-oxides cells.
Inhibition of 95% and 100% of the nitrate reductaseactivities was observed in aerobic and anaerobic wholecells respectively. This difference of azide inhibition
chain between nitrate reductase and nitrite reductase.
level on the nitrate reductase activity may suggest that
In turn, in the partial aerobic culture with cyanide,
(i) two different nitrate reductases are active according
nitrates were not consumed during the period of
to the aeration conditions or (ii) two different electron
oxygen consumption correlated with proteins synthe-
donor pathways are used for the same nitrate reduc-
sis (11 h). On the contrary, the concentration of nitrate
sharply decreased without a lag phase in the control
The inhibitory effect of cyanide on nitrate reduc-
culture. Despite this phenomenon, after complete
tion was another aspect contributing to these two
disappearance of oxygen in the test culture, the nitrate
previous hypotheses. It was demonstrated that a high
consumption was similar to that observed under anaero-
concentration of cyanide (100 µM) completely inhib-
bic conditions. During fully aerobic conditions, at 10
ited denitrification under anaerobiosis as well as under
µM, denitrification was not noticed, whereas the growth
aerobiosis (data not shown). Figures 6a and 6b show
rate was similar to that obtained in the control fully
the evolution of anions during batch culture with and
aerobic culture, implying no inhibition of oxidases.
without cyanide at 10 µM (added in the medium at time
This suggests that 10 µM of cyanide directly inhibits
0) under anaerobic (a) and partial aerobic (b) condi-
the aerobic denitrifying enzymes. Thus, cyanide influ-
tions. Under anaerobic conditions, nitrate consump-
ences denitrification under aerobic conditions by to-
tion, with or without cyanide, followed the same
tally inhibiting the nitrate reduction, whereas under
pattern during 50 h. In contrast, a higher quantity of
anaerobic conditions, no effect was observed on nitrate
nitrite was accumulated in the presence of the respira-
tory inhibitor compared with the control culture. Thus,it seems that a concentration of 10 µM of cyanide has
Effect of triton X-100, azide, and DDC on nitrite
no effect on the nitrate reductase synthesized under
reductase activity. Figure 4 shows that triton partially
anaerobic conditions. However, it partially inhibits the
inhibited anaerobic nitrate consumption because of its
nitrite reductase or an intermediate of the respiratory
effect on membrane disorganization. In the same time,
no nitrite was accumulated in the medium, whereas in
served with Pseudomonas aeruginosa, a classical
the control culture the nitrite concentration increased
oxygen-sensitive denitrifier [12]. With Comamonas
sharply to reach a peak of 80 mg N-NO2 · L21. By
sp., the decrease in nitrate reduction rate was corre-
disturbing the permeability of the cytoplasmic mem-
lated with a decrease in nitrogen gas production and
brane, triton could act on the antiport system nitrate/
partial nitrite, nitric oxide, and nitrous oxide accumula-
nitrite by preventing the nitrate from joining the active
tion. However, the presence of these denitrifying
site of the enzyme and by holding the nitrite in the
intermediates during continuous culture, at a dissolved
cytoplasm. On the contrary, triton, when added in
oxygen concentration of 100% of air saturation (data
batch culture with nitrite as final electron acceptor, had
not shown), implies that the four denitrifying enzymes
no direct effect on nitrite reduction whatever the
are active and synthesized under aerobic conditions.
culture and preculture conditions used. This may be
The same conclusions are drawn, since higher aerobic
owing to periplasmic localization of the nitrite reduc-
nitrate–nitrite reduction rates were observed in culture
without chloramphenicol compared with that obtained
Azide and DDC were chosen to show the possible
with the protein synthesis inhibitor (Figs. 1–3).
existence of a copper nitrite reductase or copper
These aerobic nitrate and nitrite reductase activi-
intermediates as pseudoazurin in the respiratory chain:
ties could not be interpreted in terms of assimilation
the former reacts with ferric centers, the latter links to
because (i) (NH4)2SO4, used as the nitrogen source,
copper centers. Nitrite reduction seems to be insensi-
classically inhibits nitrate assimilation, (ii) ammonium
tive to respiratory inhibitors during anaerobic batch
disappearance was well correlated with biomass pro-
assay with nitrate as the final electron acceptor:
duction, and (iii) during continuous culture under
addition of azide or DDC resulted on one hand in total
oxygen-saturated conditions, nitrogen was produced,
inhibition of nitrate reductase activity (Fig. 5) and, on
which is the direct demonstration of denitrification
the other hand, in a complete disappearance of nitrite
reaction. Van Niel and associates [38], using nitrate
accumulated during the previous hours. In the same
labeled on nitrogen, have confirmed the idea of
way, nitrate reductase activity measured by enzymatic
Robertson and Kuenen [27] that a complete denitrify-
assay on whole cells is expressed as the quantity of
ing system is present under aerobic conditions in T.
nitrate disappearing because no nitrite accumulated
pantotropha. According to the experiments done with
during the test. In the presence of the different
chloramphenicol on cells never subjected to nitrate,
inhibitors, batch assays with nitrite as the final electron
the existence of a constitutive nitrate reductase was
acceptor ended up with the same conclusion: whatever
proposed to explain the possible aerobic denitrification
the culture and preculture conditions, azide and DDC
in Comamonas sp. An opposite conclusion resulted
had no effect on nitrite reductase activity.
from the same experiments made with Paracoccusdenitrificans (NCIB 8944): nitrite, nitric oxide, nitrousoxide, and nitrogen gas were not produced during
Discussion
anaerobic and partial aerobic cultures with chloram-
Previous work had generally underlined that in many
phenicol. Without the protein inhibitor, anaerobic
bacteria, synthesis and activity of denitrifying en-
denitrification started with a long lag period of 11 h,
zymes could not occur under aerobiosis. Our work
corresponding to the synthesis of a de novo nitrate
leads to a modified conclusion, and the results allow us
to propose a scheme to explain the nitrate-oxygen
The close relation between the denitrifying en-
co-respiration in Comamonas sp. strain SGLY2. The
zymes and the electron transport pathway, the different
first experiments done with the strain demonstrated its
kinds of nitrate transport, the genetic and the regula-
ability to use simultaneously the two electron accep-
tory system of synthesis, and the activity of the
tors [22]. Aeration of the culture resulted in a decrease
enzymes are now well studied [11]. These enzymes are
in the nitrate reduction rate: 1.85 µmol NO 2
shown to work in vitro in the presence of oxygen [1, 3,
mg21 of protein under anaerobic culture against 0.287
24]. However, in vivo, other aspects have to be
in aerated culture. These values lie close to that found
considered to explain the possible aerobic denitrifica-
with Thiosphaera pantotropha, in which the rate of
tion. From a bioenergetic point of view, the idea of
acetate-dependent nitrate reduction is around 1.6 µmol
co-respiration seems illogical, because energy produc-
· min21 · mg21 of protein at dissolved oxygen
tion is higher with oxygen than nitrate and because the
concentration less than 30%, and 0.8 at 30–80% of air
main regulatory factor of denitrification is the redox
saturation [29]. In contrast, at a concentration of
potential of the respiratory chain [17]. For example,
0.25% of air saturation, no denitrification was ob-
the presence of oxygen in a P. denitrificans culture
D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.
implies preferential diversion of electrons to oxygen
tory effect of DDC was noticed on both anaerobic and
owing to the modification of the redox potential of the
aerobic nitrite reductase activities. On the other hand, a
coupler ubiquinol/ubiquinone [9, 10]. Hernandez and
large amount of nitrous oxide was accumulated in the
colleagues [13] have also shown that, using Pseudomo-
gaseous phase, perhaps owing to the blocking of a
nas aeruginosa, oxygen indirectly inhibits enzyme
multi-copper nitrous oxide reductase. Moreover, no
activities by oxidizing the key molecules of the
nitrite accumulation was observed during aerobic
antiport nitrate-nitrite system. Moreover, it is known
culture on nitrate with azide. These observations
that oxygen regulates nitrate respiration by suppress-
suggest the existence of a cdl-type nitrite reductase.
ing enzyme synthesis: a FnR-like binding site, re-
According to the experiments done with and without
quired for anaerobic gene expression in Escherichia
protein inhibitor, this inducible enzyme seems to be
coli, is present in cells of P. denitrificans [11, 33] and
active and synthesized under aerobic conditions. At
Pseudomonas stutzeri [8]. Bell et al. [2] have reported
this point, the behavior of the strain differs from that of
that T. pantotropha uses a periplasmic nitrate reductase
T. pantotropha: using polyclonal antibodies, Moir [20]
while denitrifying aerobically and employs another
has shown that the cdl-type nitrite reductase was not
membrane-bound reductase for anaerobic denitrifica-
expressed under aerobic conditions. The status of
tion. Thus, the oxygen inhibitory effect on nitrate-
aerobic denitrifier of T. pantotropha is then not clear.
nitrite antiport system is evaded. The diversion of the
In fact, it has been shown that there is a close
electron flow to the denitrifying enzymes was ex-
relationship between T. pantotropha and Paracoccus
plained by the hypothesis of the ‘‘bottleneck’’ [26]. denitrificans [19, 35]. Moreover, although a periplas-
Using sodium azide as respiratory inhibitor, Van Niel
mic nitrate reductase seems to be synthesized in
and coworkers [38] showed that nitrogen gas produc-
Paracoccus denitrificans [32], no aerobic denitrifica-
tion stopped immediately after the addition of 10 mM
tion was noticed in this strain. Kuenen and Robertson
azide to the aerobic cell suspension. Conversely, 0.02
[18], in their last experiments, observed that the
mM azide is just enough to inhibit the nitrate reductase
aerobic denitrification rate of T. pantotropha is now
synthesized under anaerobic conditions [23]. In the
equivalent to 5% of that found under anaerobic
same way, the different levels of triton and azide
conditions versus 50% at the beginning of their
inhibition between aerobic and anaerobic cells suggest
the existence of two nitrate reductases in SGLY2: one
Physiological observations on Comamonas sp.
‘‘aerobic’’ enzyme insensitive to membrane damage
strain SGLY2, using different respiratory or protein
caused by triton and less sensitive to azide, and one
synthesis inhibitors and detergent, explain the ability
‘‘anaerobic’’ enzyme partially inhibited by 0.02% of
of the strain to denitrify under aerobic conditions.
From an ecological point of view, existence of this
Cyanide effect is another factor contributing to the
kind of aerobic denitrifier is interesting to explain
hypothesis of existence of two nitrate reductases. The
nitrogen losses of agronomic system. It has thus to be
hypothesis of existence of two electron donor path-
considered in agricultural practices, especially for the
ways for one nitrate reductase was rejected after
mode of application of nitrogen fertilizers.
comparison of the azide effect on nitrate and nitrite
Literature Cited
reduction under anaerobic conditions. Since azide hadno effect on nitrite reductase activity, whereas it
1. Bell LC, Ferguson SJ (1991) Nitric and nitrous oxide reducta-
completely inhibited nitrate reductase activity, we can
ses are active under aerobic conditions in cells of Thiosphaerapantotropha. Biochem J 273:423–427
conclude that the nitrate reduction inhibition is a direct
2. Bell LC, Richardson DJ, Ferguson SJ (1990) Periplasmic and
effect on the enzyme and not a consequence of
membrane-bound respiratory nitrate reductases in Thiosphaera
inhibition of an intermediate of the respiratory chain. pantotropha. FEBS Microbiol Lett 265:85–87
Moreover, one enzyme could not be differently inhib-
3. Bell LC, Page MD, Berks BC, Richardson DJ, Ferguson SJ
ited by the same inhibitor. This is why existence of two
(1993) Insertion of transposon Tn5 into a structural gene of themembrane-bound nitrate reductase of Thiosphaera Pantotro-pha results in anaerobic overexpression of periplasmic nitrate
No aerobic denitrification can occur with the
reductase activity. J Gen Microbiol 139:3205–3214
existence of a copper-type nitrite reductase in the
4. Berks BC, Baratta D, Richardson DJ, Ferguson SJ (1993)
denitrifying enzymatic system because of its proper-
Purification and characterization of a nitrous oxide reductase
ties to reduce oxygen to toxic peroxides. Moir and
from Thiosphaera pantotropha. Implications for the mecha-nism of aerobic nitrous oxide reduction. Eur J Biochem
associates [21] have purified a cdl-type nitrite reduc-
tase from T. pantotropha and its presumably electron
5. Berks BC, Richardson DJ, Robinson C, Reilly A, Aplin RT,
donor pseudoazurin. With Comamonas sp., no inhibi-
Ferguson SJ (1994) Purification and characterization of the
periplasmic nitrate reductase from Thiosphaera pantotropha.
23. Richardson DJ, Ferguson SJ (1992) The influence of carbon
substrate on the activity of the periplasmic nitrate reductase in
6. Bonin P, Gilewicz M (1991). A direct demonstration of
aerobically grown Thiosphaera pantotropha. Arch Microbiol
co-respiration of oxygen and nitrogen oxides by Pseudomonasnautica: some spectral and kinetic properties of the respiratory
24. Richardson DJ, Bell LC, McEwan AG, Jackson JB, Ferguson
components. FEBS Microbiol Lett 80:183–188
SJ (1991) Cytochrome c2 is essential for electron transfer to
7. Brons HJ, Zehnder AJB (1990) Aerobic nitrate and nitrite
nitrous oxide reductase from physiological substrates in Rhodo-
reduction in continuous cultures of Escherichia coli E4. Arch
bacter capsulatus and can act as an electron donor to the
reductase in vitro: correlation with photoinhibition studies. Eur
8. Cuypers H, Zumft WG (1993) Anaerobic control of denitrifica-
tion in Pseudomonas stutzeri escapes mutagenesis of an
25. Robertson LA, Kuenen JG (1983) Thiosphaera pantotropha
FNR-like gene. J Bacteriol 175:7236–7246
gen. nov., a facultatively anaerobic, facultatively autotrophic
9. Ferguson SJ (1987) Denitrification: a question of the control
sulphur bacterium. J Gen Microbiol 129:2847–2855
and organization of electron and ion transport. Trends Biochem
26. Robertson LA, Kuenen JG (1984a) Aerobic denitrification: old
wine in new bottle. Antonie van Leeuwenhoek 50:525–544
10. Ferguson SJ (1992) The periplasm. In: Cole JA (ed.), Procary-
27. Robertson LA, Kuenen JG (1984b) Aerobic denitrification: a
otic structure and function, a new perpective. Society for
controversy revived. Arch Microbiol 139:351–354
General Microbiology Symposium 47, pp 297–315
28. Robertson LA, Kuenen JG (1990) Combined heterotrophic
11. Ferguson SJ (1994) Denitrification and its control. Antonie van
nitrification and aerobic denitrification in Thiosphaera panto-tropha and other bacteria. Antonie van Leeuwenhoek 57:139–
12. Hernandez D, Rowe JJ (1987) Oxygen regulation of nitrate
uptake in denitrifying Pseudomonas aeruginosa. Appl Environ
29. Robertson LA, Van Niel EWJ, Torresmans RAM, Kuenen JG
(1988) Simultaneous nitrification and denitrification in aerobic
13. Hernandez D, Dias FM, Rowe JJ (1991) Nitrate transport and
chemostat cultures of Thiosphaera pantotropha. Appl Environ
its regulation by oxygen in Pseudomonas aeruginosa. Arch
30. Robertson LA, Cornelisse R, De Vos P, Hadioetomo R, Kuenen
14. John P (1977) Aerobic and anaerobic bacterial respiration
JG (1989) Aerobic denitrification in various heterotrophic
monitored by electrodes. J Gen Microbiol 98:231–238
nitrifiers. Antonie van Leeuwenhoek 56:289–299
15. Ko¨rner H, Zumft WG (1989) Expression of denitrification
31. Sacks LE, Barker HA (1949) The influence of oxygen on
enzymes in response to the dissolved oxygen level and
nitrate and nitrite reduction. J Bacteriol 58:11–22
respiratory substrate in continuous culture of Pseudomonas
32. Sears HJ, Ferguson SJ, Richardson DJ, Spiro S (1993) The
stutzeri. Appl Environ Microbiol 55:1670–1676
identification of a periplasmic nitrate reductase in Paracoccus
16. Krul JM, Veeningen R (1977) The synthesis of the dissimila-
denitrificans. FEMS Microbiol. Lett 113:107–111
tory nitrate reductase under aerobic conditions in a number of
33. Spiro S (1992) An FNR-dependent promoter from Escherichia
denitrifying bacteria, isolated from activated sludge and drink-
coli is active and anaerobically inducible in Paracoccusdenitrificans. FEMS Microbiol Lett 98:145–148
17. Kucera I, Dadak V (1983) The effect of uncoupler on the
34. Stickland LH (1931) The reduction of nitrates by E. coli.
distribution of the electron flow between the terminal acceptors
hydrogen and nitrite in the cells of Paracoccus denitrificans.
35. Stouthamer AH (1992) Metabolic pathways in Paracoccusdenitrificans and closely related bacteria in relation to phylog-
18. Kuenen JG, Robertson LA (1994) Combined nitrification and
eny of prokariotes. Antonie van Leeuwenhoek 61:1–33
denitrification processes. FEMS Microbiol Rev 15:109–117
36. Thomas KL, Llyod D, Boddy L (1994) Effects of oxygen, pH
19. Ludwig W, Mittenhuber G, Friedrich CG (1993) Transfer of
and nitrate concentration on denitrification by PseudomonasThiosphaera pantotropha to Paracoccus denitrificans. Int J
species. FEMS Microbiol Lett 118:186
37. Thomsen JK, Iversen JJL, Cox RP (1993) Interactions between
20. Moir JWB (1993) Ph.D. thesis, University of Oxford, UK
respiration and denitrification during growth of Thiosphaera
21. Moir JWB, Baratta D, Richardson DJ, Ferguson SJ (1993) The
pantotropha in continuous culture. FEMS Microbiol Lett
purification of a cdl-tyle nitrite reductase and the absence of a
copper nitrite reductase from the aerobic denitrifier Thio-
38. Van Niel EWJ, Robertson LA, Cox RP, Kuenen JG (1992)
sphaera pantotropha; the role of pseudoazurin as an electron
Inhibition of denitrification and oxygen utilization by Thio-sphaera pantotropha. J Gen Appl Microbiol 38:553–558
22. Patureau D, Davison J, Bernet N, Moletta R (1994) Denitrifica-
39. Zumft WG (1992) The denitrifying procaryotes, p 554–582. In
tion under various aeration conditions in Comamonas sp, strain
Balows A, Tru¨per HG, Dworkin M, Harder W, Schleifer KH
(eds.), The procaryotes, 2nd ed., Springer-Verlag, Berlin
RESUME FOR PROFESSOR DR. ABUL KALAM AZAD CHOWDHURY 1. PERSONAL DETAILS: Chairman (State Minister) , University Grants Commission of Vice Chancellor , University of Dhaka, Sept’ 96 to Nov 2001. Invited Professor , 2003, Dept. of Biology, Conservatoire National des Arts et Métiers (CNAM), 75003 Paris, France. Professor, Department of Clinical Pharmacy and Pharmacology
and staff of the Michigan Ear Institute, web site at www.michiganear.com Michigan Ear Institute higan Ear Institute (248) 865-4444 phone Jack M. Kartush, MD ORS Dennis I. Bojrab, MD Michael J. LaRouere, MD John J. Zappia, MD, FACS Eric W. Sargent, MD, FACS Seilesh C. Babu, MD Eleanor Y. Chan, MD Providence Medical Building Beaumont Medical Building Oakwo
 CURRENT MICROBIOLOGY Vol. 32 (1996), pp. 25–32
Study of the Denitrifying Enzymatic System of Comamonas sp.
CURRENT MICROBIOLOGY Vol. 32 (1996), pp. 25–32
Study of the Denitrifying Enzymatic System of Comamonas sp.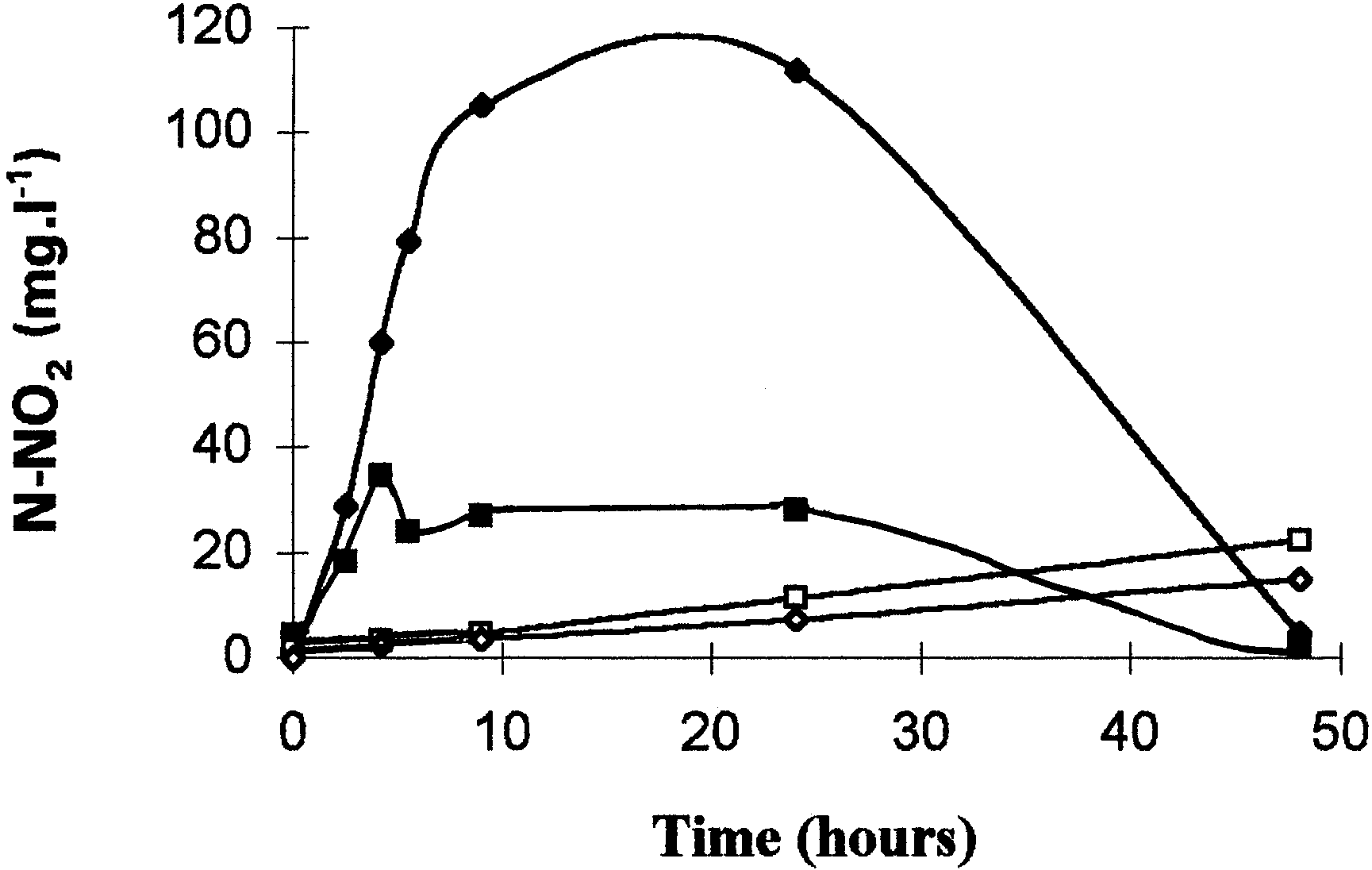
 D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.
D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.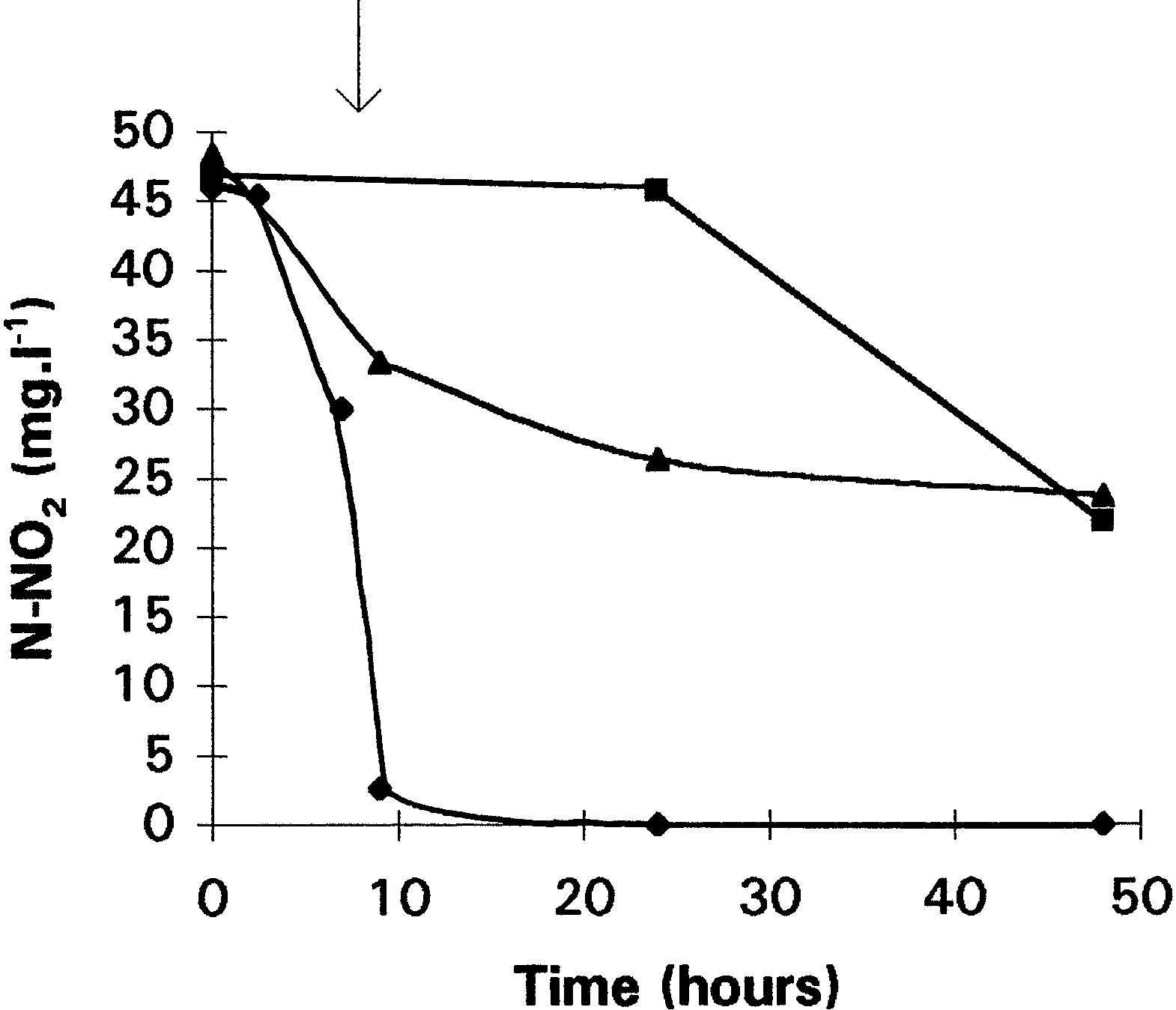
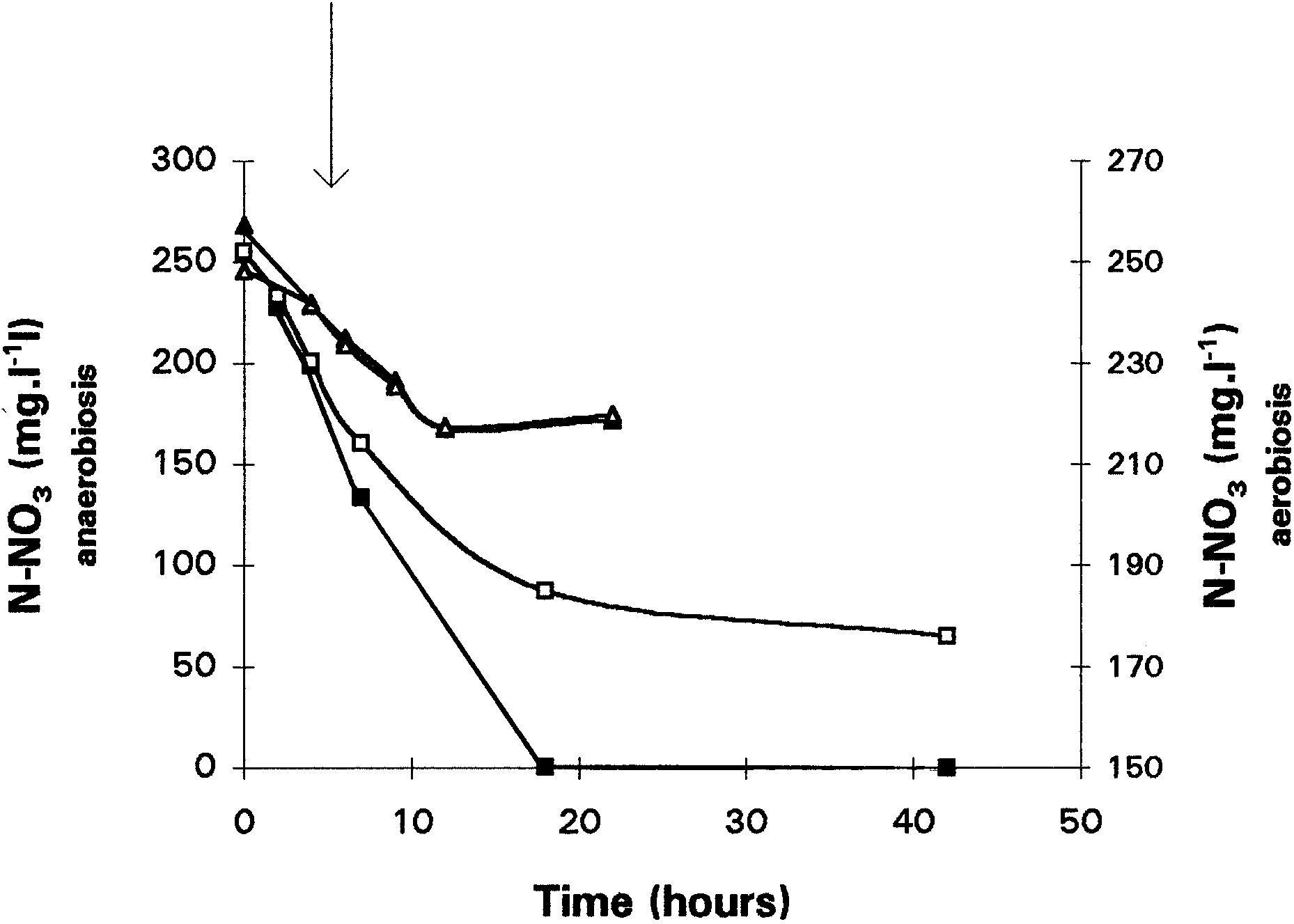
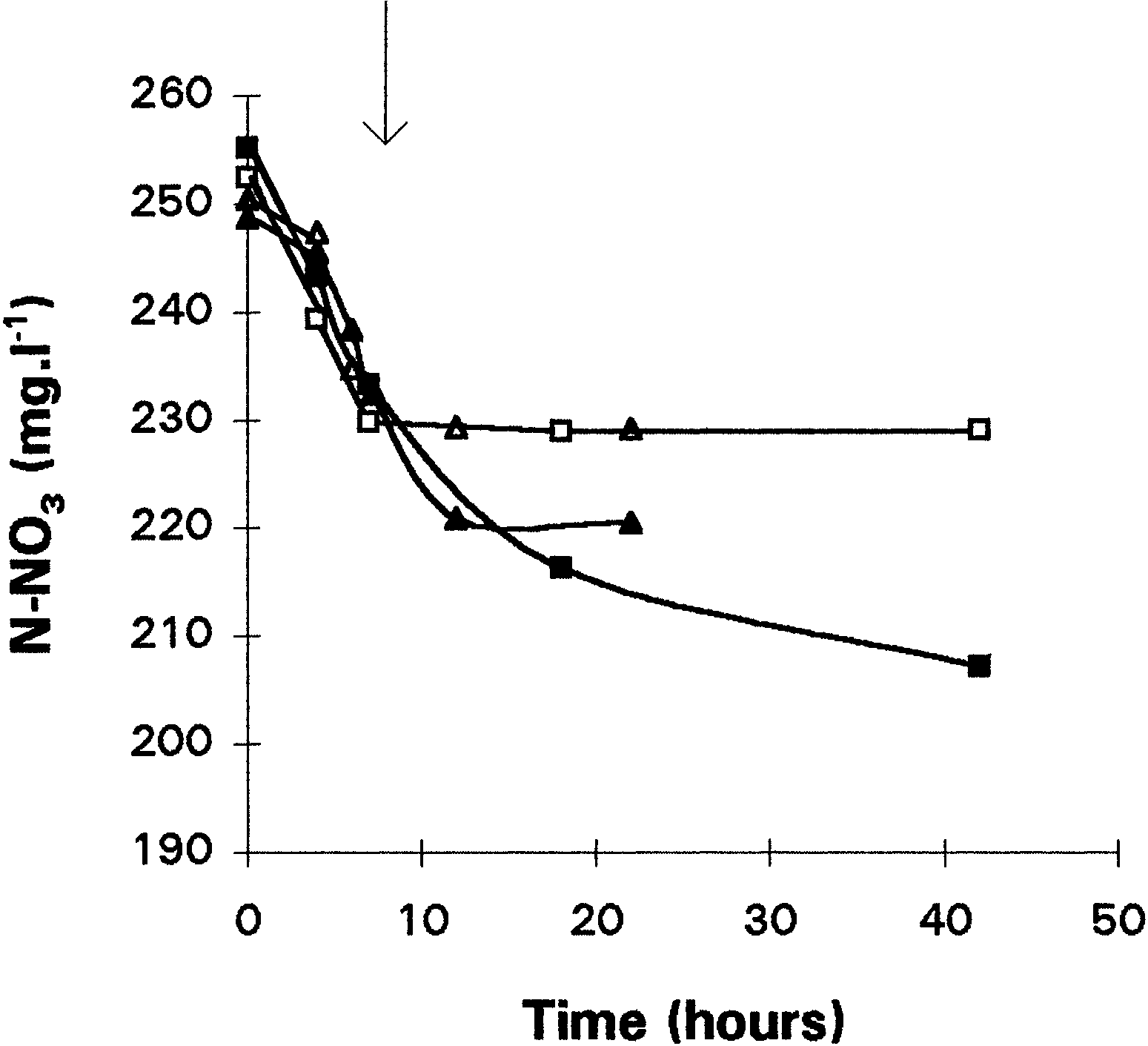 Fig. 3. Consumption of nitrite during anaerobic (M), partial aerobic
Fig. 4. Influence of triton (0.02%) on the consumption of nitrate
(U), and total aerobic cultures (Q) of Comamonas sp. The arrow
during anaerobic (M, N) and total aerobic (Q, S) cultures of
indicates the time at which oxygen has completely disappeared in
Comamonas sp. Cultures were inoculated with anaerobic cells. M,
the partial aerobic culture. Cultures were inoculated with non-
Q, control cultures; N, S; Cultures with triton; =, addition of
present in the cells. In contrast, partial or fully aerobicconsumption of nitrite started after a smaller lag period(4 h). This consumption was correlated with nitrousoxide and nitrogen production (data not shown).
Fig. 3. Consumption of nitrite during anaerobic (M), partial aerobic
Fig. 4. Influence of triton (0.02%) on the consumption of nitrate
(U), and total aerobic cultures (Q) of Comamonas sp. The arrow
during anaerobic (M, N) and total aerobic (Q, S) cultures of
indicates the time at which oxygen has completely disappeared in
Comamonas sp. Cultures were inoculated with anaerobic cells. M,
the partial aerobic culture. Cultures were inoculated with non-
Q, control cultures; N, S; Cultures with triton; =, addition of
present in the cells. In contrast, partial or fully aerobicconsumption of nitrite started after a smaller lag period(4 h). This consumption was correlated with nitrousoxide and nitrogen production (data not shown).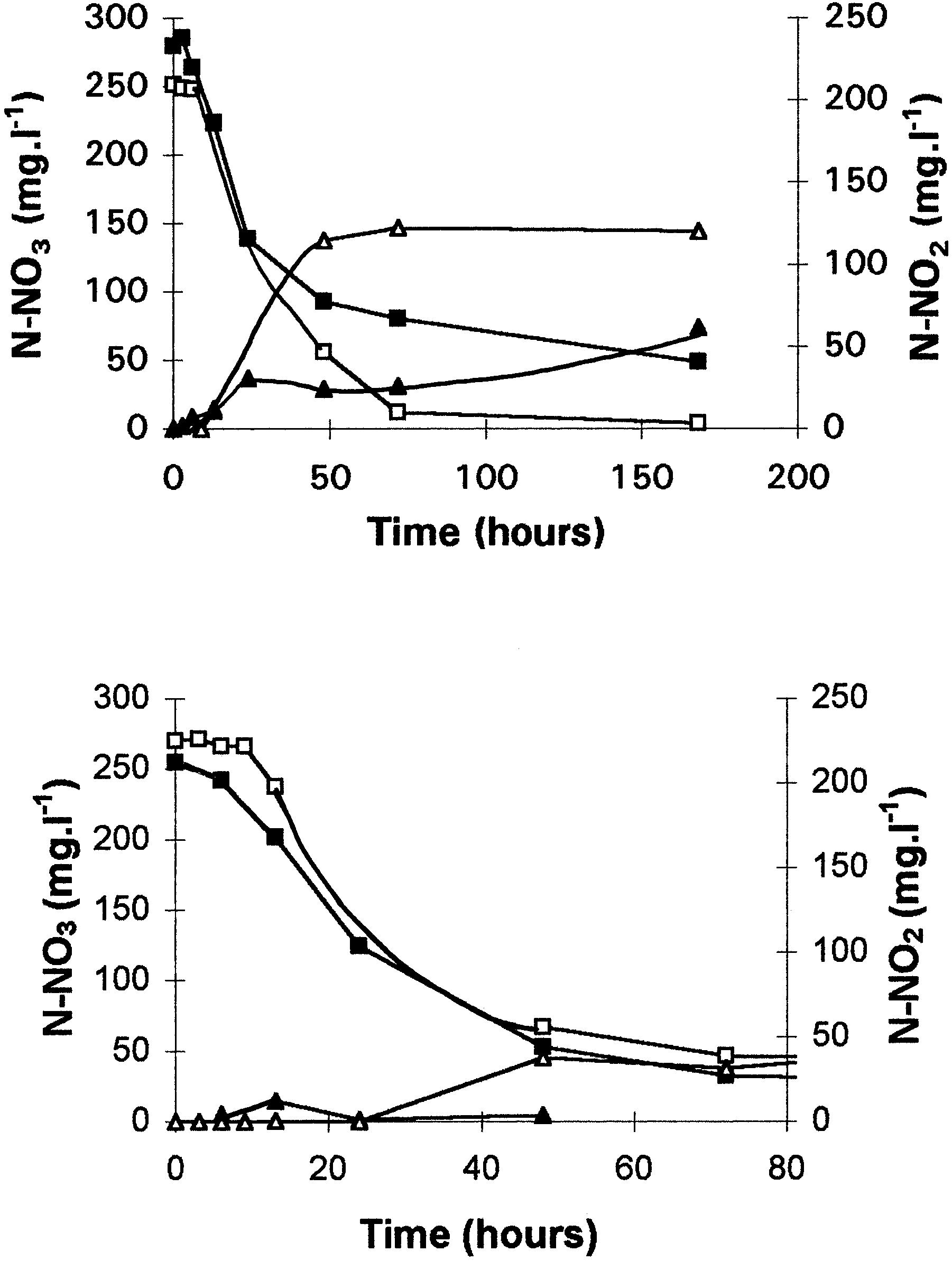 D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.
D. Patureau et al.: Denitrifying Enzymatic System of Comamonas sp.