Overexpression of the hereditary hemochromatosis protein, HFE, in
HeLa cells induces an iron-de¢cient phenotype
Barbara Corsia, Sonia Levia, Anna Cozzia, Angelo Cortia, Domenico Altimarea,
aDibit, Department of Biological and Technological Research, IRCCS H. San Ra¡aele, Via Olgettina 58, 20132 Milan, Italy
bDepartment of Biomedical Technologies, University of Brescia, Brescia, Italy
Abstract A transfectant HeLa cell clone expressing HFE under
crypt cells of the small intestine [12] and in Kup¡er cells, in
the control of a tetracycline-repressible promoter was generated.
liver and brain sinusoidal cells, and in scattered epithelial cells
HFE expression was fully repressed by the presence of
in the crypts of the small and large intestine [13].
doxycycline, while it was strongly induced by growth in the
HFE associates with the transferrin receptor (TfR) more
absence of doxycycline. HFE accumulation was accompanied by
tightly at neutral (pH 7.5) than at acidic pH (pH 6) and the
a large (V10-fold) decrease in H- and L-ferritin levels, by a
binding reduces TfR a¤nity for Fe-transferrin [14,15]. In
V3^4-fold increase in transferrin receptor, and a V2-fold
transfected cells an association between HFE and TfR occurs
increase in iron regulatory protein activity. These indices of
in the endoplasmic reticulum/cis-Golgi compartment soon
cellular iron deficiency were reversed by iron supplementation
after synthesis, it stabilizes HFE protein from degradation
complexes. The overexpressed HFE immunoprecipitated to-
[16], and reduces cellular iron uptake [17,18]. HeLa cell clones
gether with transferrin receptor, indicating a physical association
which is the likely cause for the observed V30% decrease in
overexpressing HFE showed low levels of endogenous ferritins
55Fe-transferrin incorporation after 18 h incubation. In the HFE-
and a reduced capacity to incorporate 55Fe-transferrin, while
expressing cells the reduction in transferrin-mediated iron
they took up 55Fe-NTA at the same rate as control clones
incorporation was partially compensated by a V30% increase
[17]. The pH-dependent association with TfR appears a major
in non-transferrin iron incorporation from 55Fe-NTA, evident
and important linkage of HFE with cellular iron metabolism,
after prolonged, 18 h, incubations. The findings indicate that
but its physiological role in the regulation of body iron ab-
HFE binding to transferrin receptor reduces cellular iron
sorption and tissue distribution is unclear.
availability and regulates the balance between transferrin-
We describe the production of a HeLa cell clone expressing
mediated and non-transferrin-mediated cellular iron incorpora-
HFE under the control of a tetracycline-inducible promoter.
Analysis of the clone showed that HFE overexpression is
z 1999 Federation of European Biochemical Societies.
accompanied by a large (V10-fold) decrease of H- and
Key words: Hemochromatosis; Iron metabolism;
L-ferritin accumulation, by a V3^4-fold increase in TfR pro-
Recombinant protein; HFE protein; Transferrin receptor
tein and by a strong activation of iron responsive element
(IRE) binding activity of the iron regulatory proteins (IRPs)
which are the typical indices of cellular iron de¢ciency. The
cells have a reduced capacity to incorporate 55Fe-transferrin,
while they have an increased capacity to accumulate non-
Hereditary hemochromatosis (HHC) is a common autoso-
mal recessive disorder characterized by an upregulated iron
absorption [1]. The hemochromatosis gene encodes the HFE
protein (formerly HLA-H) resembling major histocompatibil-
2.1. DNA, expression and puri¢cation of HFE protein
ity complex class I molecules [2]. The HFE Cys282CTyr
Full length human HFE cDNA was generated by RT-PCR from
mutation, which is homozygous in s 70% of HHC patients
mRNA extracted from human pulmonary cells. The cDNA was sub-
cloned into pCDNA3.1 vector (Invitrogen) and pUDH10-3 vector
[3^5], results in the loss of a structural disul¢de bond, prevents
(Clontech) fused to the myc-tag peptide at the C-terminus. The
association with L2-microglobulin (L2m) and proper presenta-
cDNA sequence encoding HFE ectodomains (residues 26^304) was
tion to cell surfaces [6,7]. Mice deleted for the HFE or the
subcloned into vector pET12b for expression in Escherichia coli, fused
L2m gene show a fast accumulation of iron in the parenchyma
to a polyhistidine tag. DNA sequencing con¢rmed that the plasmids
encoded the correct protein sequence reported in [2]. Transformation
cells of liver similar to that observed in HHC [8,9]. The arti-
of E. coli strain BL21(DE3)pLysS and protein expression were per-
¢cially induced C282Y mutation in mice produces an HHC-
formed essentially as described [19]. HFE was puri¢ed by a¤nity
like phenotype less severe than that of HFE null mice [10].
chromatography on Ni-NTA Sepharose, and protein puri¢cation
HFE mRNA is expressed in various tissues with the highest
was con¢rmed by SDS-PAGE and Coomassie blue staining. Protein
levels in liver and intestine [2], and immunohistochemical
concentration was determined with the BCA assay (Pierce) calibrated
HFE was found in the syncytiotrophoblasts of human placen-
ta [11], in the epithelial cells of the alimentary tract, in the
2.2. Antibodies, metabolic labeling and SDS-PAGE
Mouse anti-HFE antibodies were elicited by the puri¢ed and dena-
tured recombinant HFE protein from E. coli. Antibody speci¢city was
assessed by Western blotting and immunoprecipitation. To increase
*Corresponding author. Fax: (39) (2) 2643-4844.
antibody production we induced antibody-containing ascitic £uid in
some mice by intraperitoneal injection of pristane [20]. Anti-myc-tag
0014-5793 / 99 / $20.00 ß 1999 Federation of European Biochemical Societies. All rights reserved.
PII: S 0 0 1 4 - 5 7 9 3 ( 9 9 ) 0 1 3 3 0 - 7
B. Corsi et al./FEBS Letters 460 (1999) 149^152
antibody 9E10 was purchased from Sigma and anti-transferrin recep-
tor antibody from Zimed, anti-ferritin H- and L-chain monoclonal
antibodies and ELISA assays for H- and L-ferritins calibrated with
recombinant H- and L-ferritin homopolymers are described in [21].
Metabolic labeling with [35S]methionine, immunoprecipitations, SDS-
PAGE and £uorography were performed as described in [19]. Immu-
nocytochemistry was performed on ¢xed cells permeabilized with
0.5% Triton X-100 using anti-myc-tag antibody (2 Wg/ml) followed
by TRITC anti-mouse IgG rabbit antibodies (Dako) and the stain
visualized by £uorescence microscopy.
HeLa-Tet-O¡ cells (Clontech) were co-transfected with plasmid
pUDH10-3-HFE and with plasmid pTK-Hyg (Clontech) using the
calcium phosphate method [19]. The colonies selected with hygro-
mycin D (150 Wg/ml) and doxycycline (2 Wg/ml) were screened for
the HFE gene by PCR and for HFE expression by immunoprecipita-
tion of metabolically labeled cells with anti-myc antibody 9E10. The
Fig. 1. Time course of HFE expression in HeLa transfected cells.
selected cells were maintained in Dulbecco's modi¢ed Eagle's medium
Soluble lysates (30 Wg) of transfected cells grown in 2 Wg/ml doxycy-
(DMEM, Gibco) supplemented with 10% fetal bovine serum (Clon-
cline (+) or in the absence of doxycycline for the indicated days
tech), 100 Wg/ml G418 (Geneticin, Sigma), 150 Wg/ml hygromycin D
were subjected to SDS-PAGE, transferred to nitrocellulose and de-
(Clontech) 100 U/ml penicillin, 100 Wg/ml streptomycin, 1 mM L-glu-
tected with mouse anti-HFE antibody (1:4000), anti-transferrin re-
tamine and with (Dox+) or without (Dox3) 2 Wg/ml doxycycline
ceptor antibody (1:1000) or anti-H-ferritin monoclonal antibody
rHO2 (7 Wg/ml) and the appropriate peroxidase-labeled secondary
antibody (1:100). HFE and TfR were separated on 12% polyacryl-
amide gels under reducing conditions, while ferritins were separated
Subcon£uent HFE-HeLa-Tet-O¡ cells were lysed and 30 Wg of
on 7.5% polyacrylamide gels under non-denaturing conditions. The
soluble protein loaded on SDS-PAGE. For HFE and TfR analysis,
mobility of the 45^47 kDa bands of HFE, of the V94 kDa band
the samples were denatured by heating at 100³C for 10 min in the
of TfR and of the ferritin bands is indicated by arrows. The ELI-
presence of 2% 2-mercaptoethanol, and run in 12% polyacrylamide
SA-derived cellular concentrations of H- and L-ferritins on the indi-
gels. For ferritin analysis, the samples were not heated to avoid pro-
cated days of induction are shown at the bottom. MW: molecular
tein disassembly and loaded on 7.5% polyacrylamide gels. After trans-
weight standards. Representative data from two independent experi-
fer, the nitrocellulose ¢lters were incubated with anti-HFE antibody
(dilution 1:4000), with anti-TfR antibody (dilution 1:1000), or with
anti-H ferritin rHO2 monoclonal antibody (concentration 7 Wg/ml)
followed by secondary, peroxidase-labeled antibody (Envision,
Dako). Bound activity was revealed by ECL (Amersham).
obtained with the same antibody when the clone was main-
tained in the presence of 2 Wg/ml doxycycline (Dox+) (not
Bandshift experiments for IRP activity were performed as in [22].
shown). Immunocytochemistry staining with the anti-myc
Cells (2U105) subjected to various treatments were lysed and 2 Wg
samples of soluble protein were incubated with a molar excess of 32P-
antibody produced a strong, mainly intracellular decoration
labeled H-ferritin IRE probe, RNase T1 and heparin in the presence
in the Dox3 transfected cells and no signi¢cant background
or absence of 2% 2-mercaptoethanol [22]. RNA protein complexes
in Dox+ cells (not shown). These ¢ndings indicated that the
were separated on 6% non-denaturing PAGE and exposed to auto-
HeLa clone (HFE-HeLa-Tet) expresses a myc-tagged HFE
functional in binding TfR and that the expression is fully re-
Cells (2U105) were grown for various lengths of time in the pres-
The time course of HFE accumulation after doxycycline
ence of 2 WCi/ml of 55Fe-NTA (0.4 WM Fe(III), 4 WM NTA) in
removal was analyzed by Western blotting. We used mouse
DMEM, 0.5% fetal calf serum, 0.5% bovine serum albumin, 150 WM
antisera elicited by the denatured recombinant HFE produced
ascorbate. The cells were washed and lysed in 0.3 ml lysis bu¡er
(20 mM Tris-HCl pH 8.0, 200 mM LiCl, 1 mM EDTA, 0.5%
in E. coli, which is speci¢c for HFE in immunoblotting and
NP40), centrifuged and 10 Wl samples of the soluble fraction were
does not cross-react with HLA molecules (not shown). The
mixed with 0.5 ml of Ultima Gold (Packard) and counted for 3 min
antibody did not detect HFE in the homogenate of Dox3
in a scintillator counter (Packard). In other experiments the cells were
cells at days 1^3 and recognized the 45^47 kDa HFE doublet
incubated with 0.5 WM 55Fe-transferrin for 18 h in serum-free me-
from day 7 on (Fig. 1). From densitometry of the blots in
dium, and treated and counted as above.
comparison with those of known amounts of recombinant
HFE from E. coli we estimated an HFE level of about
500 ng/mg of total soluble proteins which remained constant
from day 7 to 15. Staining of the blots with an anti-TfR
To study the e¡ect of HFE expression on cellular iron me-
antibody which recognizes denatured TfR from SDS-PAGE
tabolism we generated stable HeLa transfectants with the
[17] revealed the expected V94 kDa band in all cell homoge-
HFE gene under the control of a tetracycline-responsive pro-
nates with an intensity that increased with the time of growth
moter. The screening for HFE expression was based on im-
in Dox3 to about 3^4-fold over Dox+ at day 7 (Fig. 1). An
munoprecipitation of [35S]methionine-labeled cells with the
anti-H-ferritin monoclonal antibody was used to stain blot-
9E10 antibody speci¢c for the myc-tag peptide attached at
tings from non-denaturing PAGE and it detected the typical
the C-terminus of the protein. From a clone grown in the
slow moving band of assembled ferritin (V500 kDa) the in-
absence of doxycycline (Dox3) we immunoprecipitated a
tensity of which steadily declined in the Dox3 cells to become
double band of 45^47 kDa attributed to the glycosylated
virtually undetectable from day 7 on (Fig. 1). For quantitative
and unglycosylated form of HFE, and a band of V94 kDa
data, the H- and L-ferritin concentrations were evaluated with
corresponding to TfR. No detectable immunoprecipitate was
ELISA assays based on speci¢c monoclonal antibodies and
B. Corsi et al./FEBS Letters 460 (1999) 149^152
recombinant H- and L-ferritin homopolymers, they showed
an about 10-fold decrease of both H- and L-ferritin in the
Dox3 cells at day 7 (Fig. 1, bottom). The ferritin concentra-
tion of Dox+ cells (250 and 27 Wg/mg for H- and L-ferritins,
respectively) was remarkably similar to that of untransfected
parent cells (250 þ 5 and 25 þ 3 Wg/mg for H- and L-ferritins,
respectively). In addition, as a control, we analyzed a di¡erent
transfected HeLa cell clone which did not express detectable
HFE protein to ¢nd that the ferritin levels were una¡ected by
growth in the presence or the absence of 2 Wg/ml doxycycline
Next, we analyzed IRP activity by bandshift experiments on
cell homogenates using 32P-labeled H-ferritin IRE. In trans-
fected control HeLa cells non-expressing HFE the IRP activ-
ity was analogous in the Dox+ or Dox3 conditions (Fig. 2,
lanes 1 and 2), while in the transfected cells expressing HFE,
V2-fold higher activity was observed at day 7 in Dox3 con-
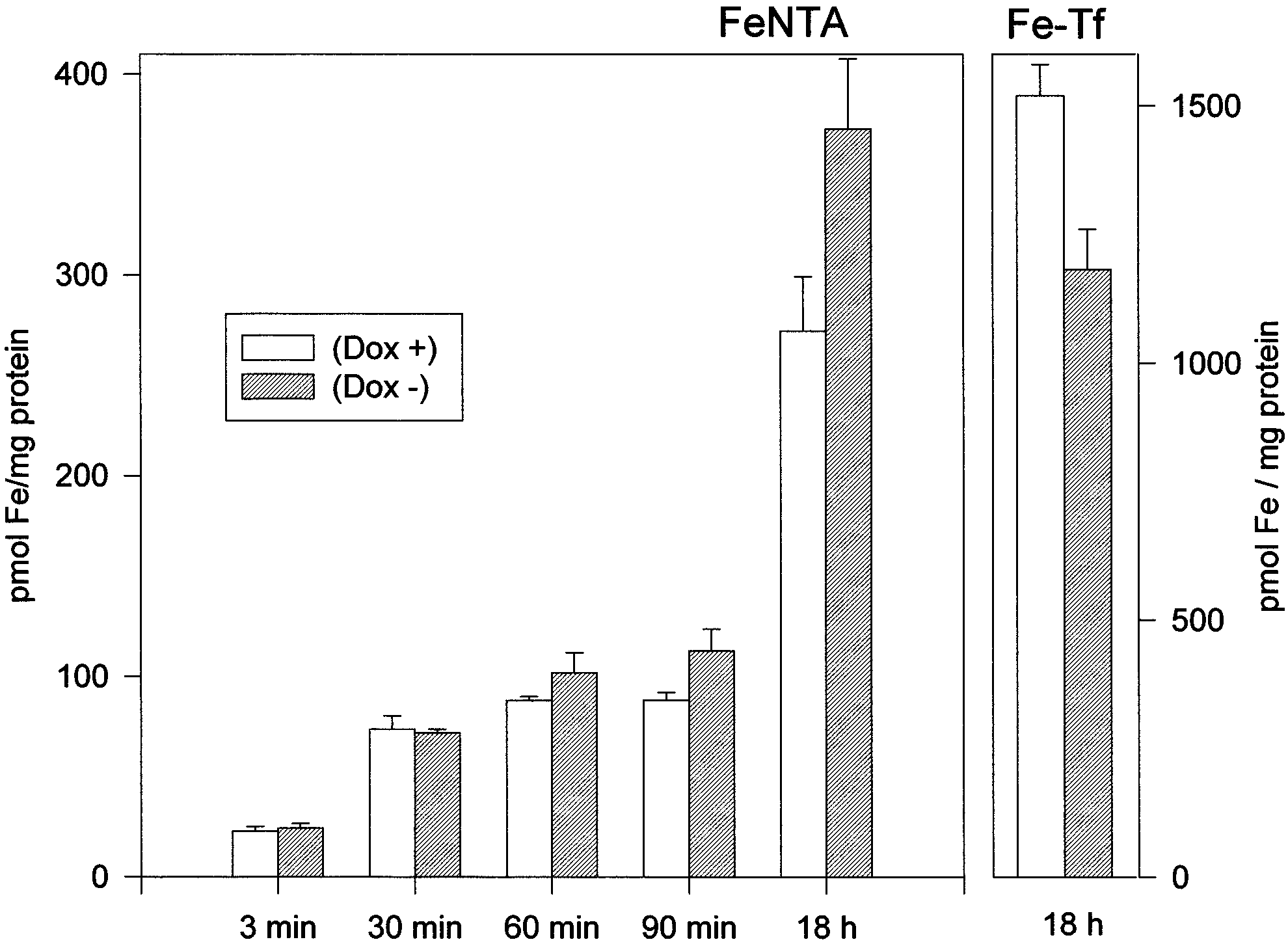
Fig. 3. HFE alters transferrin-mediated and non-transferrin-medi-
ditions (compare lanes 3 and 6 of Fig. 2). Addition of iron
ated iron incorporation after prolonged incubations. Uninduced
salts reduced IRP activity in Dox3 and Dox+ cells, while
(Dox+, empty bars) and induced transfectant cells grown 7 days in
treatment with desferrioxamine (DFO) resulted in an evident
the absence of doxycycline (Dox3, dashed bars) were incubated for
IRP activation in Dox+ cells, but not in Dox3 cells. The total
the indicated time with 0.4 WM 55Fe-NTA (Fe:NTA 1:10 molar ra-
tio, 2 WCi/ml) or for 18 h with 0.5 WM 55Fe-transferrin. Cells were
IRP activity detected after incubation with 2% 2-mercapto-
washed, lysed and the soluble extracts counted for radioactivity and
ethanol remained unchanged in the Dox+ and Dox3 cells.
analyzed for protein concentration. Data are means+S.D. of an ex-
Parallel ELISA analysis showed that FAC supplementation
periment in triplicate representative of four experiments without sig-
increased H-ferritin levels to 470 ng/mg in Dox+ and to 440
ng/mg in Dox3 cells, while DFO treatment reduced H-ferritin
levels to 6 50 ng/mg in Dox+ and Dox3 cells.
Finally, we studied cellular iron incorporation by incubat-
ing the cells with 0.5 WM 55Fe-transferrin or with 0.4 WM
HFE accumulation in our HeLa transfectant clone became
55Fe-NTA and analyzing 55Fe in the soluble extracts of the
evident after 7 days of growth in the absence of doxycycline,
cell homogenates. Dox3 cells, at day 7, incorporated V30%
probably the time needed to eliminate the excess of the drug
less 55Fe-transferrin than the Dox+ cells, after 18 h incubation
which represses the Tet promoter. The amount of accumu-
(Fig. 3), while they incorporated about 30% more iron from
lated HFE remained constant for at least a week and was
55Fe-NTA at 18 h incubation. It should be noted that iron
rather high, comparable to that of the endogenous transferrin
incorporation from 55Fe-NTA in the ¢rst 60 min of incuba-
receptor and of H-ferritin. The HFE accumulation was ac-
tion was analogous in the Dox+ and Dox3 cells, suggesting
companied by a V3^4-fold increase of TfR levels and by a
similar rates of iron incorporation.
V10-fold decrease in H- and L-ferritin levels, in agreement
with recent data [17]. IRP proteins are considered the major
sensors of cellular iron availability: their IRE binding activity
is activated by iron de¢ciency and repressed by iron availabil-
ity, although other factors such as cellular redox state and
reactive oxygen species a¡ect their functionality [23]. We ob-
served that in the conditions of maximum expression of HFE,
i.e. at day 7 of induction, IRE binding activity of IRPs was
about 2-fold higher in the HFE+ than in HFE3 cells. This
was not due to a direct e¡ect of the drug, since IRP activity in
control cells non-expressing HFE was una¡ected by doxycy-
cline (Fig. 2). Iron supplementation decreased IRP activity on
HFE+ as in HFE3 cells and increased H-ferritin content to
analogous levels. DFO treatment increased IRP activity in
HFE3, but not in the HFE+ cells in which the activity was
Fig. 2. IRP activity in the induced and non-induced cells. Autora-
diograms of representative RNA bandshift assays. Lanes 1 and 2: a
already high, consistent with the observation that DFO treat-
transfected HeLa cell control clone non-expressing HFE was grown
ment and HFE overexpression induced a similar decrease in
for 7 days in the presence (Dox+) or the absence (Dox3) of 2 Wg/
ferritin accumulation. These ¢ndings support the hypothesis
ml doxycycline. Lanes 3^14: the HFE-HeLa-Tet clone was grown
that the HFE3-induced ferritin downregulation and TfR up-
for 7 days in the presence (Dox+) of the absence (Dox3) of 2 Wg/
regulation are caused by an iron-mediated e¡ect on IRP ac-
ml doxycycline, as in Fig. 1; cells were untreated (lanes C), supple-
mented with 100 WM ferric ammonium citrate (lanes FAC) or 100
tivity. Thus, HFE overexpression limits cellular iron availabil-
WM desferrioxamine for 18 h (lanes DFO). The soluble lysates (2 Wg
ity and causes an iron-de¢cient phenotype.
protein per lane) were incubated with a 32P riboprobe spanning the
The upregulation of TfR in response to iron de¢ciency is
IRE of the ferritin H-chain in the absence or presence of 2% 2-mer-
expected to increase transferrin iron uptake, while we ob-
captoethanol (L-SH) and the RNA-protein complexes separated on
non-denaturing gel electrophoresis and exposed to autoradiography.
served a signi¢cant V30% decrease, in agreement with re-
Representative data of three independent experiments.
ported data [17]. This is likely caused by the physical associ-
B. Corsi et al./FEBS Letters 460 (1999) 149^152
ation of HFE with TfR which was shown to reduce TfR
S.I., Powell, L.W., Morris, C.P. and Walsh, T.P. (1996) Nature
capacity to bind Fe-transferrin [15^18]. The e¡ect is probably
stronger with bovine transferrin which has a lower a¤nity for
[4] Jouanolle, A.M., Fergelot, P., Gandon, G., Yaouanq, J., Le Gall,
J.Y. and David, V. (1997) Hum. Genet. 100, 544^547.
human TfR [24], and the impaired uptake of iron from bovine
[5] Carella, M., D'Ambrosio, L., Totaro, A., Grifa, A., Valentino,
transferrin, which is the major source of iron in the medium,
M.A., Piperno, A., Girelli, D., Roetto, A., Franco, B., Gasparini,
should be the cause of cellular iron de¢ciency.
P. and Camaschella, C. (1997) Am. J. Hum. Genet. 60, 828^832.
A novel ¢nding of this work is that HFE overexpression in
[6] Feder, J.N., Penny, D.M., Irrinki, A., Lee, V.K., Lebron, J.A.,
Watson, N., Tsuchihashi, Z., Sigal, E., Bjorkman, P.J. and
HeLa cells increases iron accumulation from non-transferrin-
Schatzman, R.C. (1998) Proc. Natl. Acad. Sci. USA 95, 1472^
bound iron (Fe-NTA) after long incubations. Previous studies
showed that Fe-NTA incorporation in HeLa cells is unaf-
[7] Waheed, A., Parkkila, S., Zhou, X.Y., Tomatsu, S., Tsuchihashi,
fected by cellular growth rate and DFO-induced iron deple-
Z., Feder, J.N., Schatzman, C., Britton, R.S., Bacon, B.R. and
tion, while it is increased by pretreatment with iron salts [25].
Sly, W.S. (1997) Proc. Natl. Acad. Sci. USA 94, 12384^12389.
[8] Santos, M., Schilham, M.W., Rademakers, L.H., Marx, J.J., de
However, cell lines with defective transferrin-mediated iron
Sousa, M. and Clevers, H. (1996) J. Exp. Med. 184, 1975^1985.
incorporation showed higher accumulation of 59Fe-NTA
[9] Zhou, X.Y., Tomatsu, S., Fleming, R.E., Parkkila, S., Waheed,
than control cells [26]. Thus, the observed higher 55Fe-NTA
A., Jiang, J., Fei, Y., Brunt, E.M., Ruddy, D.A., Prass, C.E.,
incorporation in HFE+ cells is probably secondary to the
Schatzman, R.C., O'Neill, R., Britton, R.S., Bacon, B.R. and
Sly, W.S. (1998) Proc. Natl. Acad. Sci. USA 95, 2492^2497.
partial inactivation of TfR activity.
[10] Levy, J.E., Montross, L.K., Cohen, D.E., Fleming, M.D. and
In conclusion, the present data demonstrate that HFE over-
Andrews, N.C. (1999) Blood 94, 9^11.
expression in HeLa cells reduces cellular iron availability and
[11] Parkkila, S., Waheed, A., Britton, R.S., Bacon, B.R., Zhou,
support a direct involvement of HFE in cellular iron metab-
X.Y., Tomatsu, S., Fleming, R.E. and Sly, W.S. (1997) Proc.
olism. However, their physiological signi¢cance in HHC re-
Nat. Acad. Sci. USA 94, 13198^13202.
[12] Waheed, A., Parkkila, S., Saarnio, J., Fleming, R.E., Zhou, X.Y.,
mains unclear, since they suggest that the inactivation of nat-
Tomatsu, S., Britton, R.S., Bacon, B.R. and Sly, W.S. (1999)
ural HFE (e.g. in C282Y homozygous) should favor Tf-
Proc. Natl. Acad. Sci. USA 196, 1579^1584.
mediated and decrease non-Tf-mediated tissue iron incorpo-
[13] Bastin, J.M., Jones, M., O'Callaghan, C.A., Schimanski, L., Ma-
ration. However, in HHC patients and in HFE knockout mice
son, D.Y. and Townsend, A.R. (1998) Br. J. Haematol. 103, 931^
the Tf-mediated basolateral iron uptake in duodenal crypt
[14] Feder, J.N., Tsuchihashi, Z., Irrinki, A., Lee, V.K., Mapa, F.A.,
enterocytes is more likely reduced than increased [12,27],
Morikang, E., Prass, C.E., Starnes, S.M., Wol¡, R.K., Parkkila,
while the non-Tf-mediated iron incorporation in absorptive
S., Sly, W.S. and Schatzman, R.C. (1997) J. Biol. Chem. 272,
duodenal enterocytes and probably also in parenchymal cells
of the liver and other organs is increased [1,27]. Possibly the
[15] Lebron, J.A., Bennett, M.J., Vaughn, D.E., Chirino, A.J., Snow,
P.M., Mintier, G.A., Feder, J.N. and Bjorkman, P. (1998) Cell
unphysiological high overexpression of HFE in the transfec-
tant cells may produce a paradoxical e¡ect, or HFE's role in
[16] Gross, C.N., Irrinki, A., Feder, J.N. and Enns, C.A. (1998)
iron metabolism may be more complex than the only associ-
[17] Roy, C.N., Penny, D.M. and Feder, J.N. (1999) J. Biol. Chem.
[18] Salter-Cid, S., Brunmark, A., Li, Y., Leturq, D., Peterson, P.A.,
Acknowledgements: The ¢nancial support of Telethon Italy (Grant
Jackson, M.R. and Yang, Y. (1999) Proc. Natl. Acad. Sci. USA
E.649) is gratefully acknowledged. The work was partially supported
by CNR Targeted Project in Biotechnology to P.A. and A.A.
[19] Corsi, B., Perrone, F., Bourgeois, R., Beaumont, C., Panzeri,
M.C., Cozzi, A., Sangregorio, R., Santambrogio, P., Albertini,
A., Arosio, P. and Levi, S. (1998) Biochem. J. 330, 315^320.
[20] Luo, W. and Lin, S.H. (1997) BioTechniques 123, 630^632.
[1] Edwards, C.Q., Gri¡en, L.M., Goldgar, D., Drummond, C.,
[21] Cozzi, A., Levi, S., Bazzigaluppi, E., Ruggeri, G. and Arosio, P.
Skolnick, M.H. and Kushner, J.P. (1998) New Engl. J. Med.
(1989) Chim. Clin. Acta 184, 197^206.
[22] Leibold, E.A. and Munro, H.N. (1988) Proc. Natl. Acad. Sci.
[2] Feder, J.N., Gnirke, A., Thomas, W., Tsuchihashi, Z., Ruddy,
D.A., Basava, A., Dormishian, F., Domingo, R.Jr., Ellis, M.C.,
[23] Hentze, M.W. and Kuhn, L.C. (1996) Proc. Natl. Acad. Sci.
Fullan, A., Hinton, L.M., Jones, N.L., Kimmel, B.E., Kronmal,
G.S., Lauer, P., Lee, V.K., Loeb, D.B., Mapa, F.A., McClelland,
[24] Penhallow, R.C., Brown-Mason, A. and Woodworth, R.C.
E., Meyer, N.C., Mintier, G.A., Moeller, N., Moore, T., Mor-
(1986) J. Cell Physiol. 128, 251^260.
ikang, E., Prass, C.E., Quintana, L., Starnes, S.M., Schatzman,
[25] Kaplan, J. and Jordan, I. (1991) J. Biol. Chem. 266, 2997^3004.
R.C., Brunke, K.J., Drayna, D.T., Risch, N.J., Bacon, B.R. and
[26] Sturrock, A., Alexander, J., Lamb, J., Craven, C.M. and Kaplan,
Wol¡, R.K. (1996) Nature Genet. 13, 399^408.
J. (1990) J. Biol. Chem. 265, 3139^3145.
[3] Jazwinska, E.C., Cullen, L., Bus¢eld, F., Pyper, W.R., Webb,
[27] Kuhn, L.C. (1999) Trends Biochem. Sci. 24, 164^166.
TEXAS CHILDREN WITH LYME DISEASE From the Texas Lyme Coalition Virginia, aged 15, high achieving student, vacationed in the Yucatan with her family. Eight weeks later, pediatrician extracted a tick from her arm. Two days later all family members came down with a flu-like illness: fever near 101; extreme fatigue. No rash. Three seemed to recover. Virginia's symptoms persisted: fever; headac
Press Release Zurich, 11 May 2010 Information on New Value’s portfolio company Bogar AG Bogar’s Board extended with industry expert – New Value participates in capital increase Bogar AG, a portfolio company of New Value, (SIX: NEWN), nominates to elect Peter Haensli as new Board member on the occasion of an upcoming shareholders meeting. Mr Haensli is a longtime pet in
 Overexpression of the hereditary hemochromatosis protein, HFE, in
HeLa cells induces an iron-de¢cient phenotype
Barbara Corsia, Sonia Levia, Anna Cozzia, Angelo Cortia, Domenico Altimarea,
aDibit, Department of Biological and Technological Research, IRCCS H. San Ra¡aele, Via Olgettina 58, 20132 Milan, Italy
bDepartment of Biomedical Technologies, University of Brescia, Brescia, Italy
Abstract A transfectant HeLa cell clone expressing HFE under
crypt cells of the small intestine [12] and in Kup¡er cells, in
the control of a tetracycline-repressible promoter was generated.
Overexpression of the hereditary hemochromatosis protein, HFE, in
HeLa cells induces an iron-de¢cient phenotype
Barbara Corsia, Sonia Levia, Anna Cozzia, Angelo Cortia, Domenico Altimarea,
aDibit, Department of Biological and Technological Research, IRCCS H. San Ra¡aele, Via Olgettina 58, 20132 Milan, Italy
bDepartment of Biomedical Technologies, University of Brescia, Brescia, Italy
Abstract A transfectant HeLa cell clone expressing HFE under
crypt cells of the small intestine [12] and in Kup¡er cells, in
the control of a tetracycline-repressible promoter was generated.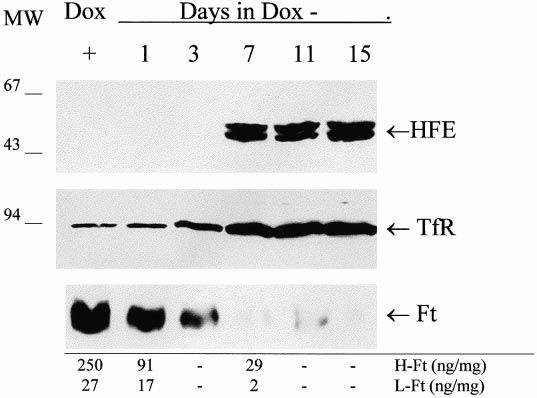 B. Corsi et al./FEBS Letters 460 (1999) 149^152
antibody 9E10 was purchased from Sigma and anti-transferrin recep-
tor antibody from Zimed, anti-ferritin H- and L-chain monoclonal
antibodies and ELISA assays for H- and L-ferritins calibrated with
recombinant H- and L-ferritin homopolymers are described in [21].
B. Corsi et al./FEBS Letters 460 (1999) 149^152
antibody 9E10 was purchased from Sigma and anti-transferrin recep-
tor antibody from Zimed, anti-ferritin H- and L-chain monoclonal
antibodies and ELISA assays for H- and L-ferritins calibrated with
recombinant H- and L-ferritin homopolymers are described in [21].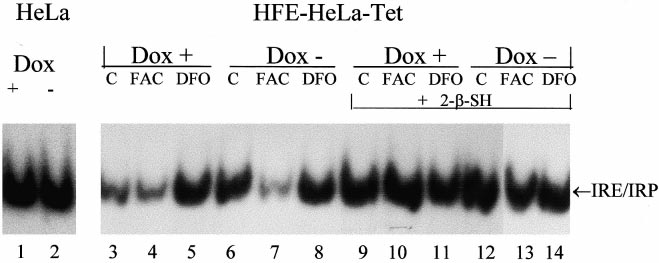
 B. Corsi et al./FEBS Letters 460 (1999) 149^152
recombinant H- and L-ferritin homopolymers, they showed
an about 10-fold decrease of both H- and L-ferritin in the
Dox3 cells at day 7 (Fig. 1, bottom). The ferritin concentra-
tion of Dox+ cells (250 and 27 Wg/mg for H- and L-ferritins,
respectively) was remarkably similar to that of untransfected
parent cells (250 þ 5 and 25 þ 3 Wg/mg for H- and L-ferritins,
respectively). In addition, as a control, we analyzed a di¡erent
transfected HeLa cell clone which did not express detectable
HFE protein to ¢nd that the ferritin levels were una¡ected by
growth in the presence or the absence of 2 Wg/ml doxycycline
Next, we analyzed IRP activity by bandshift experiments on
cell homogenates using 32P-labeled H-ferritin IRE. In trans-
fected control HeLa cells non-expressing HFE the IRP activ-
ity was analogous in the Dox+ or Dox3 conditions (Fig. 2,
lanes 1 and 2), while in the transfected cells expressing HFE,
V2-fold higher activity was observed at day 7 in Dox3 con-
Fig. 3. HFE alters transferrin-mediated and non-transferrin-medi-
ditions (compare lanes 3 and 6 of Fig. 2). Addition of iron
ated iron incorporation after prolonged incubations. Uninduced
salts reduced IRP activity in Dox3 and Dox+ cells, while
(Dox+, empty bars) and induced transfectant cells grown 7 days in
treatment with desferrioxamine (DFO) resulted in an evident
the absence of doxycycline (Dox3, dashed bars) were incubated for
IRP activation in Dox+ cells, but not in Dox3 cells. The total
the indicated time with 0.4 WM 55Fe-NTA (Fe:NTA 1:10 molar ra-
tio, 2 WCi/ml) or for 18 h with 0.5 WM 55Fe-transferrin. Cells were
IRP activity detected after incubation with 2% 2-mercapto-
washed, lysed and the soluble extracts counted for radioactivity and
ethanol remained unchanged in the Dox+ and Dox3 cells.
B. Corsi et al./FEBS Letters 460 (1999) 149^152
recombinant H- and L-ferritin homopolymers, they showed
an about 10-fold decrease of both H- and L-ferritin in the
Dox3 cells at day 7 (Fig. 1, bottom). The ferritin concentra-
tion of Dox+ cells (250 and 27 Wg/mg for H- and L-ferritins,
respectively) was remarkably similar to that of untransfected
parent cells (250 þ 5 and 25 þ 3 Wg/mg for H- and L-ferritins,
respectively). In addition, as a control, we analyzed a di¡erent
transfected HeLa cell clone which did not express detectable
HFE protein to ¢nd that the ferritin levels were una¡ected by
growth in the presence or the absence of 2 Wg/ml doxycycline
Next, we analyzed IRP activity by bandshift experiments on
cell homogenates using 32P-labeled H-ferritin IRE. In trans-
fected control HeLa cells non-expressing HFE the IRP activ-
ity was analogous in the Dox+ or Dox3 conditions (Fig. 2,
lanes 1 and 2), while in the transfected cells expressing HFE,
V2-fold higher activity was observed at day 7 in Dox3 con-
Fig. 3. HFE alters transferrin-mediated and non-transferrin-medi-
ditions (compare lanes 3 and 6 of Fig. 2). Addition of iron
ated iron incorporation after prolonged incubations. Uninduced
salts reduced IRP activity in Dox3 and Dox+ cells, while
(Dox+, empty bars) and induced transfectant cells grown 7 days in
treatment with desferrioxamine (DFO) resulted in an evident
the absence of doxycycline (Dox3, dashed bars) were incubated for
IRP activation in Dox+ cells, but not in Dox3 cells. The total
the indicated time with 0.4 WM 55Fe-NTA (Fe:NTA 1:10 molar ra-
tio, 2 WCi/ml) or for 18 h with 0.5 WM 55Fe-transferrin. Cells were
IRP activity detected after incubation with 2% 2-mercapto-
washed, lysed and the soluble extracts counted for radioactivity and
ethanol remained unchanged in the Dox+ and Dox3 cells.